


| Ресурсы VIZIT Group |




| Ресурсы VIZIT Group |


 97Модульная камера цветного изображения с объективом PINHOLEf-3,7НВЦ-80-210ННЦ-120-210(250)ННЦ-180-210(250)Оригинальный ключ VIZIT-TMРВЕ-4РВС-2РВС-4МРВС-502Рычаг-503 LONGСчитыватель RD-2Считыватель RD-4R Считыватель RD-5FУКП-12УКП-12-1УКП-12МУКП-7УКП-7МФГ-2МЭлектромеханическая защелка 54N412F
97Модульная камера цветного изображения с объективом PINHOLEf-3,7НВЦ-80-210ННЦ-120-210(250)ННЦ-180-210(250)Оригинальный ключ VIZIT-TMРВЕ-4РВС-2РВС-4МРВС-502Рычаг-503 LONGСчитыватель RD-2Считыватель RD-4R Считыватель RD-5FУКП-12УКП-12-1УКП-12МУКП-7УКП-7МФГ-2МЭлектромеханическая защелка 54N412F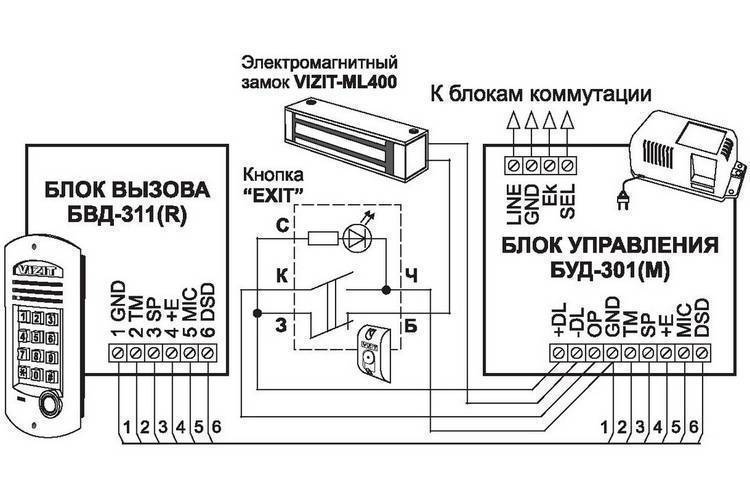 2-06VIZIT-RF2.2-10VIZIT-RF2.2-12VIZIT-TK401DVIZIT-TK401DMVIZIT-ДСН-10,5-Пр(Лв)VIZIT-ДСН-10,5-Пр(Лв)-С1VIZIT-ДСН-10,5-Пр(Лв)-С3VIZIT-ДСН-12-Пр(Лв)VIZIT-ДСН-12-Пр(Лв)-C1VIZIT-ДСН-12-Пр(Лв)-C3VIZIT-ДСН-13-Пр(Лв)VIZIT-ДСН-13-Пр(Лв)-C1VIZIT-ДСН-13-Пр(Лв)-C3VIZIT-ДСН-14-Пр(Лв)VIZIT-ДСН-14-Пр(Лв)-C1VIZIT-ДСН-14-Пр(Лв)-C3VIZIT-ДСН-15-Пр(Лв)VIZIT-ДСН-15-Пр(Лв)-C1VIZIT-ДСН-15-Пр(Лв)-C3VIZIT-ДСН-16-Пр(Лв)VIZIT-ДСН-16-Пр(Лв)-С1VIZIT-ДСН-16-Пр(Лв)-С3VIZIT-ДСНМ-10,5-Пр(Лв)VIZIT-ДСНМ-10,5-Пр(Лв)-С1VIZIT-ДСНМ-10,5-Пр(Лв)-С2VIZIT-ДСНМ-10,5-Пр(Лв)-С3VIZIT-ДСНМ-10,5-Пр(Лв)-С5VIZIT-ДСНМ-10,5-Пр(Лв)-С6VIZIT-ДСНМ-10,5-Пр(Лв)-С7VIZIT-ДСНМ-10,5-Пр(Лв)-С8VIZIT-ДСНМ-12-Пр(Лв)VIZIT-ДСНМ-12-Пр(Лв)-C1VIZIT-ДСНМ-12-Пр(Лв)-C3VIZIT-ДСНМ-13-Пр(Лв)VIZIT-ДСНМ-13-Пр(Лв)-C1VIZIT-ДСНМ-13-Пр(Лв)-C3VIZIT-ДСНМ-14-Пр(Лв)VIZIT-ДСНМ-14-Пр(Лв)-C1VIZIT-ДСНМ-14-Пр(Лв)-C3VIZIT-ДСНМ-15-Пр(Лв)VIZIT-ДСНМ-15-Пр(Лв)-C1VIZIT-ДСНМ-15-Пр(Лв)-C3VIZIT-ДСНМЦ-10,5-Пр(Лв)-С3VIZIT-ДСНМЦ-12-Пр(Лв)-C3VIZIT-ДСНМЦ-13-Пр(Лв)-C3VIZIT-ДСНМЦ-14-Пр(Лв)-C3VIZIT-ДСНМЦ-15-Пр(Лв)-C3VIZIT-ДСНТ-10,5-Пр(Лв)VIZIT-ДСНТ-10,5-Пр(Лв)-С1VIZIT-ДСНТ-10,5-Пр(Лв)-С3VIZIT-М402СMVIZIT-М430СVIZIT-М456CVIZIT-М456CMVIZIT-М457MVIZIT-МT460CMVIZIT-ПК1VIZIT-ПЛАНЕТАVPSR24-2330LIR-36-3P12БВД-101VБВД-101АБВД-104CBБВД-104VPБВД-104VPLБВД-104АБВД-107VБВД-107СБВД-2VБВД-2АБВД-311БВД-311RБВД-313RCPБВД-313RVPБВД-313TVPБВД-321БВД-321RБВД-321RCPБВД-321RVPБВД-341БВД-342FБВД-342RБВД-342R2БВД-342RTБВД-342TБВД-401AБВД-401CBБВД-401CBLБВД-401CPLБВД-401VPБВД-401VPLБВД-402CPБВД-402VPБВД-402АБВД-403VPLБВД-404CP-2БВД-404CP-4БВД-404VP-2БВД-404VP-4БВД-404А-2БВД-404А-4БВД-406CBБВД-406VBБВД-407RCBБВД-407RVBБВД-408RCB-40БВД-408RVB-40БВД-410VBLБВД-431DXKVBБВД-4AБВД-4VБВД-N100БВД-N100CPБВД-N100RБВД-N100RCPБВД-N100VPБВД-SM100RБВД-М200БВД-М200CPБВД-М200VPБВД-С100ТМБК-10БК-100БК-30БК-4БК-4VБК-4МVБПД12-1-1БПД12-2-1БУ-К100БУД-301KБУД-301МБУД-408МБУД-420Бк-30ВН-13-Пр(Лв) ВН-14-Пр(Лв)ВН-15-Пр(Лв)ВН-16-Пр(Лв)ВтулкаЗИП MVM-32NЗИП VIZIT-ML300 якорьЗИП БВД-310FCPЗИП БВД-311ЗИП БВД-311RЗИП БВД-313RЗИП БВД-313RCPЗИП БВД-313RVPЗИП БВД-313TЗИП БВД-313TCPЗИП БВД-314FЗИП БВД-314RЗИП БВД-314TЗИП БВД-321ЗИП БВД-321RЗИП БВД-342RЗИП БВД-343RЗИП БВД-432RCBЗИП БВД-N100RЗИП БВД-SM101RЗИП БК-30EЗИП БК-4MVЗИП БУД-301МЗИП КТМ-600М (600R/600F)ЗИП КТМ-602М (602R/602F)ЗИП МК-6ЗИП РВС-4КНВ-10-210(250)КНВ-20-210(250)КНВ-50-210(250)КНВ-70-210КНН-60-210(250)КНН-80-210Кабель ТРП бел.
2-06VIZIT-RF2.2-10VIZIT-RF2.2-12VIZIT-TK401DVIZIT-TK401DMVIZIT-ДСН-10,5-Пр(Лв)VIZIT-ДСН-10,5-Пр(Лв)-С1VIZIT-ДСН-10,5-Пр(Лв)-С3VIZIT-ДСН-12-Пр(Лв)VIZIT-ДСН-12-Пр(Лв)-C1VIZIT-ДСН-12-Пр(Лв)-C3VIZIT-ДСН-13-Пр(Лв)VIZIT-ДСН-13-Пр(Лв)-C1VIZIT-ДСН-13-Пр(Лв)-C3VIZIT-ДСН-14-Пр(Лв)VIZIT-ДСН-14-Пр(Лв)-C1VIZIT-ДСН-14-Пр(Лв)-C3VIZIT-ДСН-15-Пр(Лв)VIZIT-ДСН-15-Пр(Лв)-C1VIZIT-ДСН-15-Пр(Лв)-C3VIZIT-ДСН-16-Пр(Лв)VIZIT-ДСН-16-Пр(Лв)-С1VIZIT-ДСН-16-Пр(Лв)-С3VIZIT-ДСНМ-10,5-Пр(Лв)VIZIT-ДСНМ-10,5-Пр(Лв)-С1VIZIT-ДСНМ-10,5-Пр(Лв)-С2VIZIT-ДСНМ-10,5-Пр(Лв)-С3VIZIT-ДСНМ-10,5-Пр(Лв)-С5VIZIT-ДСНМ-10,5-Пр(Лв)-С6VIZIT-ДСНМ-10,5-Пр(Лв)-С7VIZIT-ДСНМ-10,5-Пр(Лв)-С8VIZIT-ДСНМ-12-Пр(Лв)VIZIT-ДСНМ-12-Пр(Лв)-C1VIZIT-ДСНМ-12-Пр(Лв)-C3VIZIT-ДСНМ-13-Пр(Лв)VIZIT-ДСНМ-13-Пр(Лв)-C1VIZIT-ДСНМ-13-Пр(Лв)-C3VIZIT-ДСНМ-14-Пр(Лв)VIZIT-ДСНМ-14-Пр(Лв)-C1VIZIT-ДСНМ-14-Пр(Лв)-C3VIZIT-ДСНМ-15-Пр(Лв)VIZIT-ДСНМ-15-Пр(Лв)-C1VIZIT-ДСНМ-15-Пр(Лв)-C3VIZIT-ДСНМЦ-10,5-Пр(Лв)-С3VIZIT-ДСНМЦ-12-Пр(Лв)-C3VIZIT-ДСНМЦ-13-Пр(Лв)-C3VIZIT-ДСНМЦ-14-Пр(Лв)-C3VIZIT-ДСНМЦ-15-Пр(Лв)-C3VIZIT-ДСНТ-10,5-Пр(Лв)VIZIT-ДСНТ-10,5-Пр(Лв)-С1VIZIT-ДСНТ-10,5-Пр(Лв)-С3VIZIT-М402СMVIZIT-М430СVIZIT-М456CVIZIT-М456CMVIZIT-М457MVIZIT-МT460CMVIZIT-ПК1VIZIT-ПЛАНЕТАVPSR24-2330LIR-36-3P12БВД-101VБВД-101АБВД-104CBБВД-104VPБВД-104VPLБВД-104АБВД-107VБВД-107СБВД-2VБВД-2АБВД-311БВД-311RБВД-313RCPБВД-313RVPБВД-313TVPБВД-321БВД-321RБВД-321RCPБВД-321RVPБВД-341БВД-342FБВД-342RБВД-342R2БВД-342RTБВД-342TБВД-401AБВД-401CBБВД-401CBLБВД-401CPLБВД-401VPБВД-401VPLБВД-402CPБВД-402VPБВД-402АБВД-403VPLБВД-404CP-2БВД-404CP-4БВД-404VP-2БВД-404VP-4БВД-404А-2БВД-404А-4БВД-406CBБВД-406VBБВД-407RCBБВД-407RVBБВД-408RCB-40БВД-408RVB-40БВД-410VBLБВД-431DXKVBБВД-4AБВД-4VБВД-N100БВД-N100CPБВД-N100RБВД-N100RCPБВД-N100VPБВД-SM100RБВД-М200БВД-М200CPБВД-М200VPБВД-С100ТМБК-10БК-100БК-30БК-4БК-4VБК-4МVБПД12-1-1БПД12-2-1БУ-К100БУД-301KБУД-301МБУД-408МБУД-420Бк-30ВН-13-Пр(Лв) ВН-14-Пр(Лв)ВН-15-Пр(Лв)ВН-16-Пр(Лв)ВтулкаЗИП MVM-32NЗИП VIZIT-ML300 якорьЗИП БВД-310FCPЗИП БВД-311ЗИП БВД-311RЗИП БВД-313RЗИП БВД-313RCPЗИП БВД-313RVPЗИП БВД-313TЗИП БВД-313TCPЗИП БВД-314FЗИП БВД-314RЗИП БВД-314TЗИП БВД-321ЗИП БВД-321RЗИП БВД-342RЗИП БВД-343RЗИП БВД-432RCBЗИП БВД-N100RЗИП БВД-SM101RЗИП БК-30EЗИП БК-4MVЗИП БУД-301МЗИП КТМ-600М (600R/600F)ЗИП КТМ-602М (602R/602F)ЗИП МК-6ЗИП РВС-4КНВ-10-210(250)КНВ-20-210(250)КНВ-50-210(250)КНВ-70-210КНН-60-210(250)КНН-80-210Кабель ТРП бел. 2×0,40МК-300-40-GRAYМК-300-40-САМОДЕЛЬ 505РВС-4РМ15-2-3РТ1×2Считыватель RD-3Считыватель RD-4TУКП-101УКП-11УКП-4УКП-8SMУКП-9МФГ2-xx-xx-xxХG-TFT817 2.5″
2×0,40МК-300-40-GRAYМК-300-40-САМОДЕЛЬ 505РВС-4РМ15-2-3РТ1×2Считыватель RD-3Считыватель RD-4TУКП-101УКП-11УКП-4УКП-8SMУКП-9МФГ2-xx-xx-xxХG-TFT817 2.5″Домофоны и видеодомофоны VIZIT. Системы контроля доступа. Металлические двери. ООО НПФ «МОДУС-Н»
Выберите изделие снятое с производстваAP-1250 SGHAP-14 SGAP-14 SGHAP-143GBS-1BS-2BS-4BS-8DL 401/55 PZ72 RtDL 428 «DOORLOCK»EXIT 300Eff-Eff 1705 E4 1GX ПанельKING NSK1633KING NSK612KING NSK633KING NSK650LCD модуль 3,5» LCD модуль 4″MC ML400TMC VISITOR-111MC VISITOR-112CMC VISITOR-112CMMC VISITOR-112MMC VISITOR-401MC VISITOR-401CMC VISITOR-401SMC VISITOR-402MC VISITOR-402CMC VISITOR-402CMMC VISITOR-402MMC VISITOR-402SMK-321MK-408-1MK-408-4-1MK-408-4-2MK-408-8MK-T30MP-440ODC-701-3.6PX ПанельRD-3RD-4FRD-4FRT-4TSA03010029VIZIT-6MVIZIT-A400VIZIT-DC503SVIZIT-K100VIZIT-K8VIZIT-KTM-40VIZIT-KTM600FVIZIT-KTM602FVIZIT-M112CVIZIT-M112CMVIZIT-M112MVIZIT-M112SVIZIT-M404CVIZIT-M404CМVIZIT-M406VIZIT-M407MVIZIT-M427CVIZIT-M440CVIZIT-M440CMVIZIT-M457МGVIZIT-M467MGVIZIT-M467MSVIZIT-ML300-40VIZIT-ML300-50VIZIT-ML400-40VIZIT-ML400-50VIZIT-ML400SVIZIT-RF2. 2-06VIZIT-RF2.2-08VIZIT-RF2.2-10VIZIT-RF2.2-12VIZIT-TK401DVIZIT-TK401DMVIZIT-ДСН-10,5-Пр(Лв)VIZIT-ДСН-10,5-Пр(Лв)-С1VIZIT-ДСН-10,5-Пр(Лв)-С3VIZIT-ДСН-12-Пр(Лв)VIZIT-ДСН-12-Пр(Лв)-C1VIZIT-ДСН-12-Пр(Лв)-C3VIZIT-ДСН-13-Пр(Лв)VIZIT-ДСН-13-Пр(Лв)-C1VIZIT-ДСН-13-Пр(Лв)-C3VIZIT-ДСН-14-Пр(Лв)VIZIT-ДСН-14-Пр(Лв)-C1VIZIT-ДСН-14-Пр(Лв)-C3VIZIT-ДСН-15-Пр(Лв)VIZIT-ДСН-15-Пр(Лв)-C1VIZIT-ДСН-15-Пр(Лв)-C3VIZIT-ДСН-16-Пр(Лв)VIZIT-ДСН-16-Пр(Лв)-С1VIZIT-ДСН-16-Пр(Лв)-С3VIZIT-ДСНМ-10,5-Пр(Лв)VIZIT-ДСНМ-10,5-Пр(Лв)-С1VIZIT-ДСНМ-10,5-Пр(Лв)-С3VIZIT-ДСНМ-10,5-Пр(Лв)-С5VIZIT-ДСНМ-10,5-Пр(Лв)-С6VIZIT-ДСНМ-10,5-Пр(Лв)-С7VIZIT-ДСНМ-10,5-Пр(Лв)-С8VIZIT-ДСНМ-12-Пр(Лв)VIZIT-ДСНМ-12-Пр(Лв)-C1VIZIT-ДСНМ-12-Пр(Лв)-C3VIZIT-ДСНМ-13-Пр(Лв)VIZIT-ДСНМ-13-Пр(Лв)-C1VIZIT-ДСНМ-13-Пр(Лв)-C3VIZIT-ДСНМ-14-Пр(Лв)VIZIT-ДСНМ-14-Пр(Лв)-C1VIZIT-ДСНМ-14-Пр(Лв)-C3VIZIT-ДСНМ-15-Пр(Лв)VIZIT-ДСНМ-15-Пр(Лв)-C1VIZIT-ДСНМ-15-Пр(Лв)-C3VIZIT-ДСНМЦ-10,5-Пр(Лв)-С1VIZIT-ДСНМЦ-12-Пр(Лв)VIZIT-ДСНМЦ-12-Пр(Лв)-C1VIZIT-ДСНМЦ-13-Пр(Лв) вставка распашнаяVIZIT-ДСНМЦ-13-Пр(Лв)-C1VIZIT-ДСНМЦ-13-Пр(Лв)-C3VIZIT-ДСНМЦ-13-Пр(Лв)-С1 вставка распашнаяVIZIT-ДСНМЦ-15-Пр(Лв) вставка распашнаяVIZIT-ДСНМЦ-15-Пр(Лв)-C1 вставка распашнаяVIZIT-ДСНТ-10,5-Пр(Лв)VIZIT-ДСНТ-10,5-Пр(Лв)-С1VIZIT-ДСНТ-10,5-Пр(Лв)-С3VIZIT-М402СVIZIT-М402СMVIZIT-М430СVIZIT-М456CVIZIT-М456CMVIZIT-М457MVIZIT-МT460CMVIZIT-ПК1VIZIT-ПЛАНЕТАБВД-101VБВД-101АБВД-104CBБВД-104VPБВД-104VPLБВД-104АБВД-107VБВД-107СБВД-2VБВД-2АБВД-311БВД-311RБВД-312RБВД-312RБВД-312TБВД-313RCPБВД-313RVPБВД-314FБВД-314FCPБВД-314RБВД-314RCPБВД-314TБВД-314TCPБВД-315FCPWБВД-316FCPWБВД-321БВД-321RБВД-321RCPБВД-321RVPБВД-341БВД-342FБВД-342RБВД-342R2БВД-342RTБВД-342TБВД-344F БВД-344RБВД-344RTБВД-401AБВД-401CBLБВД-401CPLБВД-401VPБВД-401VPLБВД-402VPБВД-403VPLБВД-404CP-2БВД-404CP-4БВД-404VP-2БВД-404VP-4БВД-404А-2БВД-404А-4БВД-406CBБВД-406VBБВД-407RCBБВД-407RVBБВД-408RCBБВД-408RCB-40БВД-408RVB-40БВД-410VBLБВД-431DXKVBБВД-444CP-1/RБВД-444CP-2/FБВД-444CP-2/RБВД-444CP-4/FБВД-444CP-4/RБВД-4AБВД-4VБВД-N100БВД-N100CPБВД-N100RБВД-N100RCPБВД-N100VPБВД-SM100БВД-SM100RБВД-SM101RБВД-SM101RCPLБВД-М200БВД-М200CPБВД-М200VPБВД-С100ТМБК-10БК-100БК-30БК-4БК-4VБК-4МVБКМ-440БПД12-1-1БПД12-2-1БУ-К100БУД-301KБУД-301МБУД-302K-80БУД-302К-20БУД-302МБУД-408МБУД-420БУД-420МБУД-420РБУД-430БУД-430БУД-430MБУД-430SБУД-485Бк-30ВН-13-Пр(Лв)ВН-14-Пр(Лв)ВН-15-Пр(Лв)ВН-16-Пр(Лв)ЗИП MVM-32NЗИП БВД-311ЗИП БВД-311RЗИП БВД-312RЗИП БВД-313RЗИП БВД-313RCPЗИП БВД-313RVPЗИП БВД-313TЗИП БВД-314FЗИП БВД-314RЗИП БВД-314TЗИП БВД-321RЗИП БВД-341ЗИП БВД-342RЗИП БВД-342RTЗИП БВД-342TЗИП БВД-343RЗИП БВД-343RCPLЗИП БВД-344RЗИП БВД-SM100RЗИП БВД-SM101RЗИП БВД-SM101TЗИП БК-30EЗИП БК-4MVЗИП БУД-301КЗИП БУД-301МЗИП МК-6ЗИП МУ-430ЗИП МУ-456ЗИП МУ-460ЗИП РВС-4AVКНВ-10-210(250)КНВ-20-210(250)КНВ-50-210(250)КНВ-70-210КНН-60-210(250)КНН-60-280КНН-80-210Кабель КДСНМКабель плоскийМК-300-40-CAМК-300-40-GRAYМОДЕЛЬ 503МОДЕЛЬ 505РВС-4РМ15-2-4РТ1×2РычагТелекамера VPSR24-2330LIR-36-3P12УКП-101УКП-11УКП-4УКП-8SMУКП-9МФГ-2-xx-yy-zzФГ-2Т-xx-yy-zz 2-06VIZIT-RF2.2-08VIZIT-RF2.2-10VIZIT-RF2.2-12VIZIT-TK401DVIZIT-TK401DMVIZIT-ДСН-10,5-Пр(Лв)VIZIT-ДСН-10,5-Пр(Лв)-С1VIZIT-ДСН-10,5-Пр(Лв)-С3VIZIT-ДСН-12-Пр(Лв)VIZIT-ДСН-12-Пр(Лв)-C1VIZIT-ДСН-12-Пр(Лв)-C3VIZIT-ДСН-13-Пр(Лв)VIZIT-ДСН-13-Пр(Лв)-C1VIZIT-ДСН-13-Пр(Лв)-C3VIZIT-ДСН-14-Пр(Лв)VIZIT-ДСН-14-Пр(Лв)-C1VIZIT-ДСН-14-Пр(Лв)-C3VIZIT-ДСН-15-Пр(Лв)VIZIT-ДСН-15-Пр(Лв)-C1VIZIT-ДСН-15-Пр(Лв)-C3VIZIT-ДСН-16-Пр(Лв)VIZIT-ДСН-16-Пр(Лв)-С1VIZIT-ДСН-16-Пр(Лв)-С3VIZIT-ДСНМ-10,5-Пр(Лв)VIZIT-ДСНМ-10,5-Пр(Лв)-С1VIZIT-ДСНМ-10,5-Пр(Лв)-С3VIZIT-ДСНМ-10,5-Пр(Лв)-С5VIZIT-ДСНМ-10,5-Пр(Лв)-С6VIZIT-ДСНМ-10,5-Пр(Лв)-С7VIZIT-ДСНМ-10,5-Пр(Лв)-С8VIZIT-ДСНМ-12-Пр(Лв)VIZIT-ДСНМ-12-Пр(Лв)-C1VIZIT-ДСНМ-12-Пр(Лв)-C3VIZIT-ДСНМ-13-Пр(Лв)VIZIT-ДСНМ-13-Пр(Лв)-C1VIZIT-ДСНМ-13-Пр(Лв)-C3VIZIT-ДСНМ-14-Пр(Лв)VIZIT-ДСНМ-14-Пр(Лв)-C1VIZIT-ДСНМ-14-Пр(Лв)-C3VIZIT-ДСНМ-15-Пр(Лв)VIZIT-ДСНМ-15-Пр(Лв)-C1VIZIT-ДСНМ-15-Пр(Лв)-C3VIZIT-ДСНМЦ-10,5-Пр(Лв)-С1VIZIT-ДСНМЦ-12-Пр(Лв)VIZIT-ДСНМЦ-12-Пр(Лв)-C1VIZIT-ДСНМЦ-13-Пр(Лв) вставка распашнаяVIZIT-ДСНМЦ-13-Пр(Лв)-C1VIZIT-ДСНМЦ-13-Пр(Лв)-C3VIZIT-ДСНМЦ-13-Пр(Лв)-С1 вставка распашнаяVIZIT-ДСНМЦ-15-Пр(Лв) вставка распашнаяVIZIT-ДСНМЦ-15-Пр(Лв)-C1 вставка распашнаяVIZIT-ДСНТ-10,5-Пр(Лв)VIZIT-ДСНТ-10,5-Пр(Лв)-С1VIZIT-ДСНТ-10,5-Пр(Лв)-С3VIZIT-М402СVIZIT-М402СMVIZIT-М430СVIZIT-М456CVIZIT-М456CMVIZIT-М457MVIZIT-МT460CMVIZIT-ПК1VIZIT-ПЛАНЕТАБВД-101VБВД-101АБВД-104CBБВД-104VPБВД-104VPLБВД-104АБВД-107VБВД-107СБВД-2VБВД-2АБВД-311БВД-311RБВД-312RБВД-312RБВД-312TБВД-313RCPБВД-313RVPБВД-314FБВД-314FCPБВД-314RБВД-314RCPБВД-314TБВД-314TCPБВД-315FCPWБВД-316FCPWБВД-321БВД-321RБВД-321RCPБВД-321RVPБВД-341БВД-342FБВД-342RБВД-342R2БВД-342RTБВД-342TБВД-344F БВД-344RБВД-344RTБВД-401AБВД-401CBLБВД-401CPLБВД-401VPБВД-401VPLБВД-402VPБВД-403VPLБВД-404CP-2БВД-404CP-4БВД-404VP-2БВД-404VP-4БВД-404А-2БВД-404А-4БВД-406CBБВД-406VBБВД-407RCBБВД-407RVBБВД-408RCBБВД-408RCB-40БВД-408RVB-40БВД-410VBLБВД-431DXKVBБВД-444CP-1/RБВД-444CP-2/FБВД-444CP-2/RБВД-444CP-4/FБВД-444CP-4/RБВД-4AБВД-4VБВД-N100БВД-N100CPБВД-N100RБВД-N100RCPБВД-N100VPБВД-SM100БВД-SM100RБВД-SM101RБВД-SM101RCPLБВД-М200БВД-М200CPБВД-М200VPБВД-С100ТМБК-10БК-100БК-30БК-4БК-4VБК-4МVБКМ-440БПД12-1-1БПД12-2-1БУ-К100БУД-301KБУД-301МБУД-302K-80БУД-302К-20БУД-302МБУД-408МБУД-420БУД-420МБУД-420РБУД-430БУД-430БУД-430MБУД-430SБУД-485Бк-30ВН-13-Пр(Лв)ВН-14-Пр(Лв)ВН-15-Пр(Лв)ВН-16-Пр(Лв)ЗИП MVM-32NЗИП БВД-311ЗИП БВД-311RЗИП БВД-312RЗИП БВД-313RЗИП БВД-313RCPЗИП БВД-313RVPЗИП БВД-313TЗИП БВД-314FЗИП БВД-314RЗИП БВД-314TЗИП БВД-321RЗИП БВД-341ЗИП БВД-342RЗИП БВД-342RTЗИП БВД-342TЗИП БВД-343RЗИП БВД-343RCPLЗИП БВД-344RЗИП БВД-SM100RЗИП БВД-SM101RЗИП БВД-SM101TЗИП БК-30EЗИП БК-4MVЗИП БУД-301КЗИП БУД-301МЗИП МК-6ЗИП МУ-430ЗИП МУ-456ЗИП МУ-460ЗИП РВС-4AVКНВ-10-210(250)КНВ-20-210(250)КНВ-50-210(250)КНВ-70-210КНН-60-210(250)КНН-60-280КНН-80-210Кабель КДСНМКабель плоскийМК-300-40-CAМК-300-40-GRAYМОДЕЛЬ 503МОДЕЛЬ 505РВС-4РМ15-2-4РТ1×2РычагТелекамера VPSR24-2330LIR-36-3P12УКП-101УКП-11УКП-4УКП-8SMУКП-9МФГ-2-xx-yy-zzФГ-2Т-xx-yy-zz |
Карта сайта | DOMOFONPORTAL.
 RU | Инструкция, Commax, домофона, Каталог, домофонов, ООО, домофоны, схема Карта сайта | DOMOFONPORTAL.RU | Инструкция, Commax, домофона, Каталог, домофонов, ООО, домофоны, схема
RU | Инструкция, Commax, домофона, Каталог, домофонов, ООО, домофоны, схема Карта сайта | DOMOFONPORTAL.RU | Инструкция, Commax, домофона, Каталог, домофонов, ООО, домофоны, схема- Главная
- Новое устройство для управления на расстоянии с помощью ИК пульта — ИК DS
- Домофоны: сценарии договорных отношений
- Выбор видеокамеры для домофонной системы
- Видеодомофон. Есть ли альтернатива?
- FAQ по открытию домофонов (взлом домофона)
- Мифы о СКУД
- Проблемы доводчиков
- Интерфейсы связи для СКУД
- Применение IP-технологий в СКУД и других системах безопасности
- Замки механические и электрозамки в системах контроля и управления доступом
- Контроллер КБД-10 – электронный модуль для управления доступом в жилые и производственные помещения
- Каталог и обзор продукции оборудования марки Effeff
- Каталог домофонов Hyundai Telecom
- Как выбрать дверной доводчик
- Инструкция по занулению домофона
- Универсальный домофонный ключ
- IZ1991 — мультифункциональный электронный ключ с защитой данных (функциональный аналог DS1991 фирмы «Maxim-Dallas Semiconductor»)
- Модуль согласования MS-4 (UDVK-2.
 0)
0) - Схемы подключения многоабонентского домофона Rainmann (Laskomex) AO-3000 и его модификаций
- РД 78.36.002-99 — ТЕХНИЧЕСКИЕ СРЕДСТВА СИСТЕМ БЕЗОПАСНОСТИ ОБЪЕКТОВ. ОБОЗНАЧЕНИЯ УСЛОВНЫЕ ГРАФИЧЕСКИЕ ЭЛЕМЕНТОВ СИСТЕМ
- Особенности проектируемой переговорной (видеопереговорной) системы
- Средства и системы контроля и управления доступом
- Индивидуальный видеодомофон JSB
- Инструкция по эксплуатации контроллера CL-01
- Инструкции и схемы подключения для домофонов ТРЕК ТМ-05.1М, ТРЕК ТМ-05.2М, ТРЕК ТМ-05.3М
- Принципиальная схема Импульс-40Д
- Кодонаборная панель ТРЕК-ТМ05м
- Контроллер ключей Touch Memory СТМ510(2003)-01, СТМ510(2003)-02.
- Продукция HID. Краткое справочное пособие
- Выбор системы контроля доступа
- Прайс-листы ведущих отечественных производителей домофонов
- Каталог оборудования марки FALCON EYE за 2009 год
- Автономные системы контроля доступа: вчера, сегодня, завтра
- HID Global — Бесконтактные смарт-карты и считыватели iCLASS
- Электромеханические запирающие устройства
- Адаптер домофонных систем — Паскаль 1510
- Бесконтактные карты доступа HID
- Правила пользования бесконтактными картами.
- Новый стандарт СКУД: основные положения
- Commax WI-4C комплект для связи по сети 220 В
- Видеодомофон Commax DPV-4LH
- Каталог новых моделей домофонов компании Commax
- Видеодомофон Commax DPV-4PE2
- Цифровой домофон POLYLOCK-PRO-PX (KEYMAN-PRO-PX)
- Каталог оборудования FERMAX за 2009-2010 год
- Руководство по эксплуатации вызывных панелей домофонов AVC-125, AVC-325, AVC-355, AVC-360
- Каталог оборудования Tantos
- Универсальный доводчик дверной ABLOY OY DC335
- Каталог продукции компании Abloy
- Вызывные панели аудиодомофонов AVC-105, AVC-105V, AVC-109
- Каталог оборудования Kocom за 2010 год
- Видеодомофоны Gardi Lux
- Электромагнитные замки серии ML-194K
- Транспондер для чтения/записи T5557 (Atmel)
- U2270B (Atmel) — микросхема считывателя бесконтактных карт
- Новости от Domofonportal.ru — установи себе виджеты от Yandex
- Микросхемы радиочастотной идентификации (RFID) производства компании Atmel
- ISO/IEC 14443 «Идентификационные карты. Бесконтактные чиповые карты. Карты с малым расстоянием считывания»
- Стандарты RFID
- Доводчики Oubao
- Устройство передачи видеосигнала по витой паре УПВ-01
- Продукция компании Dorma
- Доводчик Dorma TS 68
- Каталог оборудования марки «Marshal»
- Электромагнитные замки М1-150, М1-300, М1-400, М2-300, М2-400
- Кодовые панели ПОЛИС 51, 51ТМ, 51ЕМ, 52
- Хотите, чтобы у ВАС ”забрали” ДОМОФОН?
- Управляющие компании и обслуживание домофона
- Электромагнитные замки производства ОЛЕВС
- Зануление блока вызова домофона
- Секретные коды домофона RAIKMANN
- Домофоны для многоквартирных домов в современной России
- Общие правила выполнения монтажных и пусконаладочных работ
- Домофон своими руками.
- Несколько слов о компонентах RFID компании Texas Instruments
- Микроконтроллер с программой для установки/замены в блок вызова домофона Vizit N-100
- Инструкции для домофонов COMMAX CDV-50A и COMMAX CDV-50AM
- Новый товар — ремкомплект ЗИП «Р-М200»
- Охранно-пожарные системы от Элтис. Часть 2
- Инструкция для видеодомофонов COMMAX CDV-70A и COMMAX CDV-70AM
- Новый товар — оригинальные платы управления для многоканального подъездного домофона CDD-20 производства «Цифрал»
- МС-1 модуль сопряжения координатно-матричных домофонов с видеодомофонами.
- Охранно-пожарные системы от Элтис. Часть 1
- KC-02AB модуль сопряжения цифровых домофонов с монитором DPV-4HP
- Одноабонентская вызывная аудиопанель AVC-109
- Бесконтактный считыватель KC-1C и бесконтактный считыватель с встроенным контроллером KC-1CK
- Одноабонентская вызывная аудиопанель AVC-105
- Видеодомофон. Система безопасности или дорогая игрушка?
- Хотите, чтобы Ваш дом стал настоящей крепостью?
- Реальность и будущее видеодомофона
- Домофон или система видеонаблюдения — как следует принимать решение?
- Контроллер Touch-Memory РТМ-TM-1000/4000
- Инструкция к домофонам Keyman-Monolit, Keyman-TM и контроллеру замка Keyman TM-500
- Системы автоматизации зданий. Домофония
- Каталог оборудования FERMAX
- Домофоны для элитных домов
- Каталог оборудования AIPHONE
- Переговорные и видеопереговорные системы
- Антивандальные вызывные панели МВК-322 и МВК-324
- Исполнительные устройства домофонов
- Системы контроля доступа на жилых объектах
- Видеодомофон Commax DPV-4MTN
- Безопасность в многоквартирных домах не заканчивается на установке домофона
- Коротко о замках
- Пример дипломного проекта по теме «Система домофон»
- Коротко об аудиодомофонах
- Пример проекта системы контроля доступа
- Коротко о видеодомофонах
- Домофоны – классификация, функции, эксплуатация. Часть 3
- Электромагнитные замки. Принципы работы и предназначение
- Многоквартирные домофоны Маршал CD-2255 и CD-3255 (CD-3099, CD-3159, CD-3255)
- Видеодомофон Commax DPV-4HP
- Домофоны – классификация, функции, эксплуатация. Часть 2
- Многоквартирные домофоны РТМ
- Домофоны – классификация, функции, эксплуатация. Часть 1
- Многоквартирный домофон Маршал CD-7000TM
- Видеомонитор Kocom-600B
- Микросхема электрически стираемого перепрограммируемого ПЗУ DS2430A
- Договор на техническое обслуживание домофона
- Интерком Commax CM-200, CM-201, CM-204, CM-206, CM-211
- Микросхема электрически стираемого перепрограммируемого ПЗУ КБ1446ВГ5
- Видеодомофон Kocom KVM-604
- Каталог оборудования Accordtec
- Микросхема электронного кодового ключа К1233КТ2
- Видеодомофон Kocom KVM-624
- Антивандальная вызывная панель AVC — 305
- Видеодомофоны серий Tantos TAB-880R1205 и TAB-880R1206
- Многоквартирные домофоны серии Метаком МК99.2
- Многоквартирный домофоны Метаком МК20-ТМ (Video)
- Многоквартирные домофоны серии Метаком МК20-ТМ
- Многоквартирные домофоны КС-2002 и КС-2002+К
- Видеодомофон Kocom KVM-524 и Kocom KVM-524R
- Многоквартирный домофон КС-2006
- Видеодомофон Commax DPV-4HP и его модификации
- Каталог оборудования Commax за 2008-2009 года
- Многоквартирные домофоны Страж 2Д-01 и Страж К
- Пластиковая карта PVC с RFID чипом Mifare Ultralight.
- Многоквартирные домофоны Импульс-40Д и Импульс 40Д-1
- Пластиковая карта PVC с RFID чипом HITAG™ 2
- Каталог оборудования Цифрал (Cyfral) за 2007 год
- Каталог оборудования Commax за 2007 год
- Многоквартирный домофоны Страж 2Д-02
- Принцип действия доводчиков
- Обучающий видеокурс мастера по домофонам
- Основные принципы работы iButton фирмы Dallas Semiconductor. Часть 5
- Бесконтактные карты доступа (RFID). Часть 2
- Основные принципы работы iButton фирмы Dallas Semiconductor. Часть 4
- Бесконтактные карты доступа (RFID). Часть 1
- Как выбрать дверной доводчик. Часть 2
- Основные принципы работы iButton фирмы Dallas Semiconductor. Часть 3
- Стандарты, применяемы в радиочастотной идентификации — ISO 11784 и ISO 11785
- Эллектромагнитный замок Accord-2001 ML-194S/ML-194KS
- Устройства согласования домофонных систем с городскими телефонными линиями и телефонными аппаратами
- Радиочастотная идентификация ( RFID )
- Протокол Wiegand
- Как выбрать дверной доводчик. Часть 1
- Основные принципы работы iButton фирмы Dallas Semiconductor. Часть 2
- Электронный замок с ключём-таблеткой i-Button (DS1990A)
- Matrix II. RFID считыватель 125KHz.
- Proximity считыватель EM Marine 125KHz СP-Z2B (врезной)
- Программирование домофонов серии «Метаком 2003»
- Микросхема электронного кодового ключа К1233КТ1
- Автономный контроллер Z-396 Timer
- Основные принципы работы iButton фирмы Dallas Semiconductor. Часть 1
- Программирование «Импульс ДС-ТМ»
- Кодонаборная клавиатура URMET 1105 CODE
- Двухканальная электронная система URMET 1104 TRANSIT
- Цифровой или координатный?
- Система контроля доступа URMET 1101/1102 MAGIKEY
- Приставка Пульсар 210В «ТЕЛЕДОМ»
- Пример проекта системы контроля доступа в квартире.
- Пример проекта на монтаж видеодомофона
- Психологический расчет
- Домофон многопроводный Импульс-ДС
- Предлагаем услуги по ремонту оборудования
- Многоквартирный домофон Wekta CCD 96
- Контроллер электронных ключей KTM-512
- Модульный домофон ТРЕК-ТМ05м
- Инструкция URMET 1042 EASY 2 WIRE
- Автономный контроллер доступа KEYMAN Proximity Ex-15, Proximity Int-15, считыватели Proximity Itm, Proximity Itm-T
- Принципиальные схемы коммутаторов для домофонов Импульс Д(ДС)
- Действуем по шаблону
- Каталог аудиодомофонов и устройств контроля доступа Keyman
- Контроллеры доступа STM (СЕАН)
- Многоквартирный домофон Filman
- Принципиальные схемы домофона Импульс-40Д
- Новые инструкции к контроллерам замков Z-5R, SK-501P, RD-2000S, КТМ602M(R), A20, CL-01
- Паспорта для многоквартирных домофонов КС-2002(+К), КС-2006, Крон, паспорта блоков вызова DP420-TD22 и DP420-RD24 производства Элтис
- Возможные неисправности системы ограничения доступа «Импульс – Д», «Импульс – ДС» и способы их устранения
- Регистрация в каталоге организаций
- Цифровой или стрелочный?!
- Блок вызова DP400-TDV12, DP303-TD16
- Эмулятор ключей от домофона
- Возможные неисправности системы ограничения доступа «Страж 2Д» и способы их устранения
- Инструкции к контроллерам замка ML194. 1, TR-511, Sonix S-1, принципиальные схемы к блокам вызова домофонов Элтис ЦП100 и Rainmann CD-2000
- Инструкции к контроллерам замка Цифрал ТС, Метаком ELC-T3E, ELC-T3M, ELC-T4E, ELC-T4M, RTM-2000, LC-1, ТМК-1990А
- Борьба за чистоту и порядок в доме не повод для ограничения прав жильцов
- Домофон против консьержа
- Инструкции к домофонам Элтис ЦП100ТМ-8 и ЦП300-ТК(ВК)
- Новый товар в интернет-магазине — симплексный абонентский блок РТС-01
- Новый товар в интернет-магазине — антивандальная кнопка
- Новые модули сопряжения индивидуальных видеодомофонов с многоквартирным домофоном в интернет-магазине
- Принципиальные схемы коммутаторов для домофонов Визит БК-2(A, V), БК-4(V), БК-10, БК-30, БК-100
- Новые контроллеры ключей в интернет-магазине
- Букварь ремонтника домофонов
- Принципиальные электрические схемы абонентских трубок для домофонов Визит УКП-8SM, УКП-9M, УКП-10M, УКП-11, УКП-12M
- Принципиальная электрическая схема абонентской трубки для цифрового домофона LF-8
- Новые ремонтные комплекты в интернет-магазине
- Инструкция к домофону Rainmann (Laskomex) KD-3000
- Принципиальная схема блока вызова домофона Rainmann (Laskomex) AO-3000
- Новые ремонтные комплекты в интернет-магазине
- Принципиальная электрическая схема абонентской трубки для цифрового домофона LM-8
- Инструкции к домофонам Rainmann (Laskomex) AO-2510 (CD-2510), AO-3000
- Новые ремонтные комплекты в интернет-магазине
- Новые ремонтные комплекты в интернет-магазине
- Принципиальные схемы блока вызова Метаком МК2003-ТМ и коммутатора COM.
- Принципиальные электрические схемы для домофона «Визит» БВД С100 и мониторов VIZIT серий 401, 402
- Принципиальные электрические схемы для домофонов «Визит»
- Альбом типовых решений для инсталляции оборудования «Элтис»
- Каталог оборудования «Элтис» от 2009 года
- Новый коммутатор от «Элтис» KM300-4.2
- Домофон — материал из Википедии
- Номера резистивных ключей для Цифрала
- Компанией Метаком выпущена модификация домофона MK2003.1-4E.
- Новая модель домофона от компании Метаком МК2008-RF
- Проектирование и монтаж систем охранного телевидения и домофонов. Рекомендации. Р 78.36.008-99
- Домофоны основные элементы и особенности инсталляции
- Открытие форума
- Приглашаем к участию в проекте
- Каталог
- Карта сайта
- Магазин
- Форум
- Файловый архив
- Инструкции
- Многоабонентские домофоны Цифрал
- Паспорт Цирфал 2094.1
- Паспорт Цирфал 2094М
- Паспорт контроллера замка Цифрал ТС
- Блоки вызова серии «ССD-20» («ССD-20», «ССD-20/TC», «ССD-20/V», «ССD-20/TCV»)
- Блоки вызова серии «ССD-40» («ССD – 40″,»ССD – 40/V»,»ССD – 40/ТС»,»ССD – 40/ТСV»,»ССD – 40/Р»,»ССD – 40/РV»)
- Блоки вызова серии «ССD-2094M» («ССD-2094M», «ССD-2094M/T», «ССD-2094M/V», «ССD-2094M/TV»)
- Руководство по эксплуатации «ССD-2094. 1″, «ССD-2094.1/V», «ССD-2094.1M»,»ССD-2094.1M/V», «ССD-2094.1И», «ССD-2094.1 И/V», «ССD-2094.1/Р»
- Трубка абонентская переговорная «КЛ-2»
- Трубка абонентская переговорная «КЛМ-2»
- Трубка абонентская переговорная «КЛМ-2»
- Трубка абонентская переговорная «КМ-2»
- Трубка абонентская переговорная «КМ-2»
- Трубка абонентская переговорная «КМ-2НО»
- Трубка абонентская переговорная «КС»
- Многоабонентские домофоны Визит
- Многоабонентские домофоны Элтис
- Многоабонентские домофоны Метаком
- Прочие устройства
- Многоабонентские домофоны Irkey
- Многоабонентские домофоны Крон
- Многоабонентские домофоны Кондор
- Многоабонентские домофоны Keyman
- Многоабонентские домофоны Filman
- Многоабонентские домофоны Urmet
- Многоабонентские домофоны Трек
- Многоабонентские домофоны Wekta
- Многоабонентские домофоны Протон-Импульс
- Домофоны Kocom
- Домофоны Commax
- Домофоны Activision
- Домофоны JSB
- Домофоны Tantos
- Контроллеры доступа
- Многоабонентские домофоны Raikmann
- Многоабонентские домофоны Rainmann (Laskomex)
- Многоабонентские домофоны КС
- Многоабонентские домофоны Маршал
- Многоабонентские домофоны Цифрал
- Принципиальные схемы
- Каталоги
- Проекты
- Стандарты
- Прайс-листы
- Инструкции
- Контакты
 0)
0)
 Бесконтактные чиповые карты. Карты с малым расстоянием считывания»
Бесконтактные чиповые карты. Карты с малым расстоянием считывания» Часть 2
Часть 2 Домофония
Домофония Часть 1
Часть 1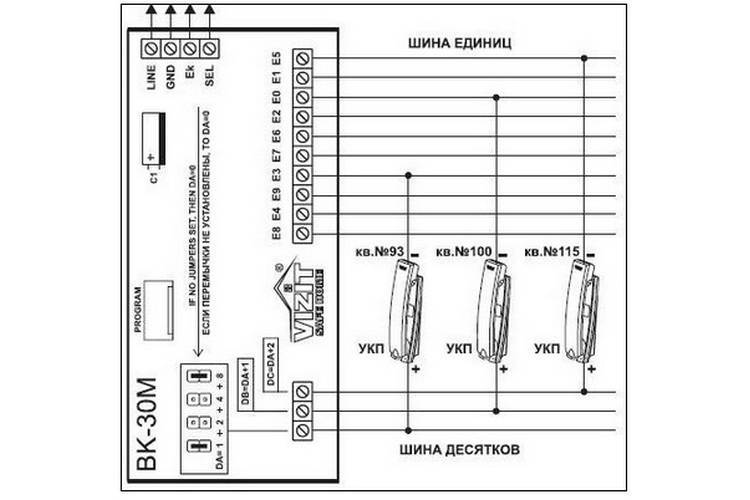
 Часть 1
Часть 1 1, TR-511, Sonix S-1, принципиальные схемы к блокам вызова домофонов Элтис ЦП100 и Rainmann CD-2000
1, TR-511, Sonix S-1, принципиальные схемы к блокам вызова домофонов Элтис ЦП100 и Rainmann CD-2000
 1″, «ССD-2094.1/V», «ССD-2094.1M»,»ССD-2094.1M/V», «ССD-2094.1И», «ССD-2094.1 И/V», «ССD-2094.1/Р»
1″, «ССD-2094.1/V», «ССD-2094.1M»,»ССD-2094.1M/V», «ССD-2094.1И», «ССD-2094.1 И/V», «ССD-2094.1/Р»Визит бвд-313 инструкция :: vscscc
14. 10.2014 09:11
Файл: Визит бвд-313 инструкция
10.2014 09:11
Файл: Визит бвд-313 инструкциявеликобритания доклад страница
волков федор григорьевич документальное видео
Описание: Блок вызова домофона БВД-313RCP.Инструкция по эксплуатации ( паспорт ) изделия, схема принципиальная приведены на странице дополнительной технической информации. Секретные коды домофонов, или как открыть домофон без ключа.Визит БВД-313 .. их можно только непосредственно прописать в замок, ручками, при . Вы искали — бвд-313r инструкция прописать ключ. R — встроенный считыватель RFID. Визит БВД-313 БВД-313Т Используется совместно с блоком управления БУД-301 или БУД-302 как составная часть многоквартирных домофонов VIZITVIZIT БВД-N100 Визит БВД-341.Avatar Vizit Домофоны Инструкция Бвд313 скачать бесплатно. Схема соединений блока вызова БВД-313x с блоком управления БУД-302М (К-20, К-80), электромагнитным замком VIZIT-ML400 и кнопкой “EXIT 300М” www.domofon.ru Инструкция по эксплуатации БВД-313x 3/4 Электромагнитный замок VIZIT-ML400 БЛОК ВЫЗОВА Многоабонентский аудиодомофон Визит VIZIT БВД-313T, блок вызова домофона БВД-313T. Блок вызова домофона БВД-313R. БВД-313 инструкция — блок вызова домофона БВД-313 инструкция.Визит — RAIKMANN инструкция — домофон Визит — RAIKMANN инструкция. В инструкции на БВД самой полезной информацией является разве что габаритные размеры и рисунки по креплению БВД, которые мало кого интересуют при наличии самого БВД перед глазами. ••• Домофон визит БВД-313R. ВЛАДИМИР ОРЛОВ Мастер (1192), закрыт.Как поменять мастер код на домофоне визит с цифр 9999 на любой другой а то шастают некоторые непрошенные гости по подъезду. Внешний вид изделия может отличаться от представленного на сайте.T — встроенный считыватель TM. руководство к действию! БВД-313Т. на кого возлагается руководство по обеспечению охраны труда. 2010) Инструкция по занулению Схема принципиальная.2013) Инструкция (ред. Инструкция (ред. Я бы не выдержал.
Блок вызова домофона БВД-313R. БВД-313 инструкция — блок вызова домофона БВД-313 инструкция.Визит — RAIKMANN инструкция — домофон Визит — RAIKMANN инструкция. В инструкции на БВД самой полезной информацией является разве что габаритные размеры и рисунки по креплению БВД, которые мало кого интересуют при наличии самого БВД перед глазами. ••• Домофон визит БВД-313R. ВЛАДИМИР ОРЛОВ Мастер (1192), закрыт.Как поменять мастер код на домофоне визит с цифр 9999 на любой другой а то шастают некоторые непрошенные гости по подъезду. Внешний вид изделия может отличаться от представленного на сайте.T — встроенный считыватель TM. руководство к действию! БВД-313Т. на кого возлагается руководство по обеспечению охраны труда. 2010) Инструкция по занулению Схема принципиальная.2013) Инструкция (ред. Инструкция (ред. Я бы не выдержал.
время и пространство документальный фильм, ваз 11174 руководство + по ремонту, вывоз мусора справка свалки r
Ваб 49 схема — samogoshka04.ru
Скачать ваб 49 схема EPUB
Опросный лист. Электрическая схема принципиальная управления выключателями В. Декларация о соответствии. Наименование параметров. Екатеринбург, ул. Русский Русский English. Высоковольтное оборудование Трансформаторно — реакторное оборудование Преобразовательная техника Проектные комплектные решения Сервисное обслуживание Испытательный центр Обучение.
Электрическая схема принципиальная управления выключателями В. Декларация о соответствии. Наименование параметров. Екатеринбург, ул. Русский Русский English. Высоковольтное оборудование Трансформаторно — реакторное оборудование Преобразовательная техника Проектные комплектные решения Сервисное обслуживание Испытательный центр Обучение.
Главная О предприятии История предприятия Награды предприятия Система качества Тендеры Корпоративная культура Карьера на предприятии Отзывы партнеров Наша продукция Высоковольтное оборудование Трансформаторно — реакторное оборудование Преобразовательная техника Проектные комплектные решения Сервисное обслуживание Испытательный центр Обучение Наши проекты Новости Публикации Фотогалерея Контакты. Выключатели применяется в одиночном исполнении. Сообщение было успешно отправлено! Разработка сайта.
Номинальный ток при постоянной во времени нагрузке, А. Номинальное напряжение постоянного тока цепей управления, В. Отключающая способность, А — в безындуктивной цепи — при индуктивности 5 — 15 мГн. Полное время отключения, с — в безындуктивной цепи — при индуктивности до 8 мГн — при индуктивности до 15 мГн. Ресурс по механической износостойкости количество включений и отключений при отсутствии тока в главной цепи , циклов.
Полное время отключения, с — в безындуктивной цепи — при индуктивности до 8 мГн — при индуктивности до 15 мГн. Ресурс по механической износостойкости количество включений и отключений при отсутствии тока в главной цепи , циклов.
Обсудить на форуме. Передача и распределение». В рамках встречи на предприятии было проведено совещание, на котором обсуждались вопросы опытной эксплуатации быстродействующих выключателей ВАБС на БАК, предложения по модернизации защитных систем постоянного тока ЦЕРН, а также рассматривались дальнейшие перспективы сотрудничества. Особое внимание представители делегации уделили осмотру участка сборки быстродействующих выключателей, где по новому заказу ЦЕРН была произведена партия дугогасительных камер для обновления уже установленных в БАК выключателей.
Во время встречи специалисты ЦЕРН высоко оценили надёжность работы установленных на БАК выключателей, большое впечатление произвела на европейских специалистов экскурсия по обновлённому производственному корпусу ВВА, гости отметили высокое качество изготовления выключателей и дугогасительных камер.
Похожее:
Устройство и работа частей БМ-302
4.1. Базовый автомобиль. В качестве базового автомобиля под размещение оборудования бурильно-крановой машины БМ-302Б использован автомобиль ГАЗ-66 повышенной проходимости.
Описание конструкции базового автомобиля изложено в эксплуатационной документации автомобиля ГАЗ-66.
4.2. Рама сварной конструкции предназначена для монтажа на ней бурильно-кранового оборудования, раздаточной коробки с лебедкой и других сборочных единиц и деталей.
В платформе автомобиля делается вырез под установку рамы, которая крепится к лонжеронам автомобиля болтами и стремянками.
4.3. Трансмиссия бурильно-крановой машины служит для передачи крутящего момента от раздаточной коробки автомобиля к вращателю бурильной штанги и к приводу барабана лебедки.
Трансмиссия состоит из коробки отбора мощности 7 (см. рис. 2), промежуточного карданного вала 8, соединяющего коробку отбора мощности с раздаточной коробкой; раздаточной коробкой 9 с лебедкой и фрикционом; промежуточного карданного вала, соединяющего раздаточную коробку с вращателем 12 бурильной штанги.
Параметры применяемых в трансмиссии шестерен и подшипников, позиции которых указаны на кинематической схеме (рис. 3), приведены в таблицах формуляра машины.
Коробка отбора мощности (рис. 4) установлена с прокладками 5 на шпильках с правой стороны раздаточной коробки автомобиля.
Привод коробки, имеющей прямозубые шестерни, осуществляется от шестерни 4 прямой передачи раздаточной коробки автомобиля.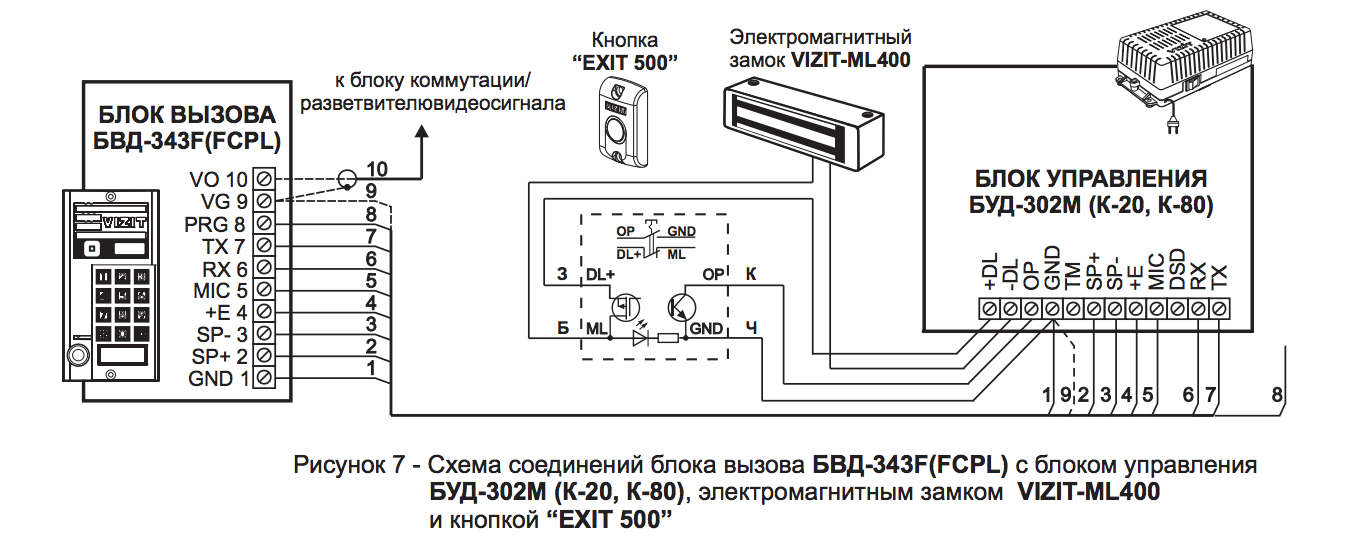 Шестерня 4 находится в постоянном зацеплении с промежуточной шестерней 3, сидящей на неподвижной оси 2.
Шестерня 4 находится в постоянном зацеплении с промежуточной шестерней 3, сидящей на неподвижной оси 2.
Передача включается при передвижении шестерни 6 по валу 1 штоком 10 с вилкой. На валу 1 установлены фланцы 7 и 9, соединенные штифтом 8, который в момент перегрузки трансмиссии срезается.
Рис. 3. Схема кинематическая принципиальная
От фланца 9 через карданный вал крутящий момент передается на раздаточную коробку.
Раздаточная коробка с лебедкой обеспечивает трехступенчатое изменение частоты вращения бура в зависимости от прочности грунта, изменение направления вращения бура и барабана лебедки.
Коробка (рис. 5, 6) смонтирована в корпусе, состоящем из картера 1, крышки 4, корпуса 6, рычага управления.
На входной вал 13 вращение от коробки отбора мощности передается карданным валом и фланцем 12. От входного вала осуществляется привод счетчика моточасов 26, и через неподвижно закрепленные шестерни 25 и 32 получает вращение вал 11.
От входного вала осуществляется привод счетчика моточасов 26, и через неподвижно закрепленные шестерни 25 и 32 получает вращение вал 11.
На шлицевом валу 11 установлены неподвижно шестерни 8, 32, 27, 28 и подвижно шестерня 9 включения лебедки. Через втулку 10 вращение передается на шестеренный насос, который крепится на фланце.
Рис. 4. Коробка отбора мощности
Рис. 5. Раздаточная коробка с лебедкой
Блок-шестерня 33 и шестерня 31 вводятся в зацепление с соответствующими шестернями вала 11, что обеспечивает три прямые передачи вращения бура; обратное вращение достигается путем ввода в зацепление шестерни 31 с шестерней 30, находящейся в постоянном зацеплении с шестерней 28.
Привод барабана 2 лебедки осуществляется зацеплением подвижной шестерни 9 с шестернями 14, 18 или 19. При этом вращение передается через вал 17 и кулачковые полу муфты 21, 22 к червячной передаче 20 и 3, на вал 5.
При этом вращение передается через вал 17 и кулачковые полу муфты 21, 22 к червячной передаче 20 и 3, на вал 5.
Кулачковая полумуфта является предохранительной и регулируется на грузоподъемность лебедки 1,25 т через пружину 23 гайками 24. Конические подшипники валов регулируются болтами 15.
Фрикцион раздаточной коробки (рис. 7) предназначен для включения привода бурильного инструмента.
Фрикцион установлен на вал шлицевым отверстием ведущего барабана 16, на наружных шлицах которого расположены ведущие диски 19. Наружный барабан 18 установлен на втулке 12, закрепленной гайкой 10.
При включении фрикциона ведущие 19 и ведомые 20 диски сжимаются пружинами 15 с помощью дисков 2 и 17, соединенных между собой пальцами 14. Выключается фрикцион при повороте рычага 6 отводки фрикциона. При этом шарики 9 выходят из лунок неподвижного кольца 8 и обоймы рычага. Перемещаясь, обойма нажимает на диск 4, он сжимает пружины 15, отводит диск 17 и освобождает диски 19 и 20. Степень нажатия дисков регулируется прокладками 11.
Выключается фрикцион при повороте рычага 6 отводки фрикциона. При этом шарики 9 выходят из лунок неподвижного кольца 8 и обоймы рычага. Перемещаясь, обойма нажимает на диск 4, он сжимает пружины 15, отводит диск 17 и освобождает диски 19 и 20. Степень нажатия дисков регулируется прокладками 11.
Рис. 6. Раздаточная коробка с лебедкой (в разрезе)
Рис. 7. Фрикцион раздаточной коробки
При выключении фрикциона одновременно происходит торможение барабана 18 тормозной колодкой 2, соединенной вилкой 3 и серьгой 1 с рычагом обоймы. Усилие прижатия тормозной колодки регулируется болтом 1.
От попадания масла в полость муфты предохраняет сальник 13.
4.4. Бурильно-крановое оборудование 2 (см. рис. 1) предназначено для передачи вращения бурильному инструменту, подачи его на забой и извлечения из скважины, а также для установки опор.
рис. 1) предназначено для передачи вращения бурильному инструменту, подачи его на забой и извлечения из скважины, а также для установки опор.
Рис. 8. Штанга бурильная
Внутри направляющей мачты монтируется бурильная штанга. К нижнему фланцу мачты крепится вращатель 12 (см. рис. 2).
Для защиты машиниста при установке опор на мачте монтируются ограждения. На оси крепления бурильного оборудования смонтирован указатель угла бурения.
Бурильная штанга (рис. 8) служит для передачи вращательного и поступательного движения бурильному инструменту. Бурильная штанга 15 перемещается по поршню 11 со штоком 13, неподвижно закрепленным в верхней части направляющей мачты. Шток с шаровой головкой 1 шарнирно крепится к корпусу 19 гайкой 17. В верхней части бурильной штанги установлен корпус 4 с втулкой 6 и уплотнениями. Рабочая жидкость подается в нижнюю полость бурильной штанги через штуцер 18 и трубу 14, а в верхнюю — через штуцер 2 и кольцевую полость штока 13. Канал 8 с клапаном предназначен для предупреждения удара гайки 10 с хвостовиком 9. Герметичность штока 13 обеспечивается резинотканевым шевронным многорядным уплотнением 5, которое при появлении течи в процессе эксплуатации подтягивается гайкой 16 через верхнее окно опорной трубы.
Рабочая жидкость подается в нижнюю полость бурильной штанги через штуцер 18 и трубу 14, а в верхнюю — через штуцер 2 и кольцевую полость штока 13. Канал 8 с клапаном предназначен для предупреждения удара гайки 10 с хвостовиком 9. Герметичность штока 13 обеспечивается резинотканевым шевронным многорядным уплотнением 5, которое при появлении течи в процессе эксплуатации подтягивается гайкой 16 через верхнее окно опорной трубы.
На бурильной штанге установлена стрелка-указатель 3 глубины бурения, которая движется по сквозному пазу мачты.
Рис. 9. Вращатель
Вращатель (рис. 9) предназначен для сообщения вращательного движения штанге и представляет собой одноступенчатый конический редуктор. Детали редуктора установлены в корпусе 4 с привернутыми к нему крышками 1, 7.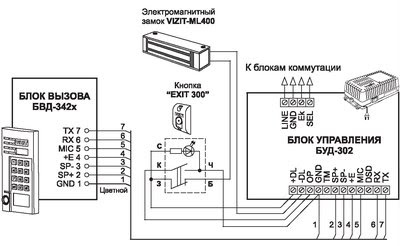 В подшипниках 2 и 6 вращается втулка 3, к которой прикреплена ведомая шестерня 5. Во втулке установлены направляющие вкладыши 12.
В подшипниках 2 и 6 вращается втулка 3, к которой прикреплена ведомая шестерня 5. Во втулке установлены направляющие вкладыши 12.
Манжеты 8 предотвращают вытекание масла из редуктора. В станке 9 помещен корпус грязеочистителя 10 со скребками 11.
Скребки прижимаются к штанге кольцом 18. Через фланец 14 вращение передается на ведущую шестерню 16, установленную в подшипниках 15 и 17. Зацепление шестерен 5 и 16 регулируется прокладками 13.
Рис. 10. Бурильный инструмент
4.5. Бурильный инструмент (рис. 10) представляет собой сменный бур, оснащенный забурником 5 и бурильными резцами, 3, которые крепятся болтами 4. Корпус 1 бура винтовой двухзаходный.
Корпус 1 бура винтовой двухзаходный.
Заслонки 2 удерживают грунт при выемке бура из скважины.
Рис. 11. Органы управления
4.6. Органы управления. Слева от машиниста расположен рычаг 5 (рис. 11) для включения и выключения коробки отбора мощности, соединенной с раздаточной коробкой автомобиля. Включать коробку следует при выключенном сцеплении автомобиля.
Для включения коробки отбора мощности надо нажать на рычаг 5 вниз, преодолевая сопротивление пружины, при этом фиксатор выйдет из паза кронштейна, и рычаг можно подать вперед в крайнее включенное положение. Привод от рычага 5 на шестерню коробки отбора мощности осуществляется через тягу 3.
С правой стороны впереди машиниста расположена педаль 6 управления подачей топлива, которая через систему блоков и каната 1 соединена с дроссельной заслонкой карбюратора двигателя автомобиля.
Впереди сиденья машиниста установлен пульт 8, где расположены органы управления гидравлической системой машины. Рычаг 14 служит для включения и выключения вращения бура, рычаг 15 — для подъема и опускания штанги, рычаг 12 — для подъема и опускания левого домкрата, рычаг 11 — для перевода бурильного оборудования из транспортного положения в рабочее и обратно, рычаг 10 — для подъема и опускания правого домкрата.
Положение рукоятки, соответствующее выполняемой операции, указано на табличках, укрепленных на панели.
Значения символов на табличках приведены на рис. 12.
На пульте управления установлен дроссель 7 (см. рис. 11), который служит для изменения расхода масла при подаче штанги для бурения. При повороте лимба против часовой стрелки расход масла увеличивается, а по часовой стрелке — уменьшается. Давление масла в гидросистеме контролируется по манометру, который включается рукояткой 13.
Запускается двигатель с рабочего места машиниста кнопкой стартера, включенного в электросхему автомобиля. Освещение приборов при работе в ночное время включается установленным на пульте управления выключателем.
Рис. 12. Таблички органов управления:
1. Включение лебедки на подъем. 2. Вращение штанги на 3-й передаче раздаточной коробки. 3. Изменение вращения штанги (реверс). 4. Вращение штанги на 1-й передаче. 5. Вращение штанги на 2-й передаче. 6. Включение лебедки на опускание. 7. Опускание штанги. 8. Подъем штанги. 9. Отключение вращения штанги. 10. Включение вращения штанги. 11. Отключение манометра. 12. Включение манометра. 13. Опускание левого домкрата. 14. Подъем левого домкрата. 15. Установка бурильного оборудования в транспортное положение. 16. Установка бурильного оборудования в рабочее положение. 17. Подъем правого домкрата. 18. Опускание правого домкрата. 19. Звуковой сигнал. 20. Регулирование подачи при бурении. 21. Включение стартера. 22. Включение освещения.
Включение освещения.
Подъем и установка опор, а также переключение скоростей раздаточной коробки осуществляются рукояткой 4, расположенной слева от сиденья машиниста. Положения рукоятки указаны в табличке, укрепленной на корпусе раздаточной коробки.
Рис. 13. Схема электрическая принципиальная
4.7 Электрооборудование. Дополнительно к электрооборудованию базового автомобиля ГАЗ-66 устанавливаются:
кнопка 1 (рис. 13) включения стартера с рабочего места машиниста, выключатель 2 для фар и ламп освещения, фара 3 освещения указателя глубины бурения и блока кранового устройства, патрон 4 с лампой 5 освещения манометра, фара 6 освещения скважины, фонарь 7 с лампой 8 контроля загрязненности фильтра, фонари 9, 11 с контрольными лампами 10, 12 давления масла в системе смазки двигателя и температуры боды в системе охлаждения двигателя, кнопка 13 включения звукового сигнала.
Источником питания указанного электрооборудования является аккумулятор и генератор базового автомобиля напряжением 12 В.
Рис. 14. Схема гидравлическая
4.8. Гидравлическая система предназначена для осуществления привода и управления механизмом установки бурильного оборудования, домкратами, бурильной штангой, фрикционом.
Гидравлическая схема представлена на рис. 14 и включает в себя: шестеренный насос 3. распределители 4 и 10, механизм установки 8 с гидрозамком 5 и дросселем 6, гидроцилиндр фрикциона 11, гидродомкраты 7, бурильную штангу 13, линейный фильтр 2, масляный бак 1, манометр 9, дроссель 12.
Управление работой гидросистемы осуществляется распределительным устройством Р80. состоящим из двух взаимосвязанных 3-золотниковых распределителей Р 80-2/2-444 и Р 80-2/3-444 клалапанно-золотникового типа.
Золотники распределителей имеют два рабочих положения и одно нейтральное. Управление золотниками осуществляется рычагами, выведенными на пульт управления.
Для защиты гидравлической системы от перегрузок в распределителе Р 80-2/2-444 установлены перепускной и предохранительный клапаны. При срабатывании предохранительного клапана, отрегулированного на давление не менее 9,8-10,8 МПа (100-110 кгс/см2), автоматически отрывается перепускной клапан, который пропускает весь поток рабочей жидкости в масляный бак.
Рис. 16. Механизм установки
Насос шестеренный НШ 50У-2 (НШ 50У-3) предназначен для нагнетания рабочей жидкости в гидросистему и состоит из корпуса 1 (рис. 15), крышки 5 и качающего узла, в который входят шестерни: ведущая 2 и ведомая 7, два компенсатора 3, левые 4 и правые 6 втулки, манжеты 8 и пластины 9.
Стык корпуса 1 с крышкой 5 уплотняется резиновым кольцом круглого сечения.
Механизм установки (рис. 16) служит для перевода бурильно-кранового оборудования в транспортное или рабочее положение, а также для удержания его в рабочем положении при бурении скважин и установке опор.
Осями 1 и 10 гидроцилиндр крепится к раме и направляющей мачте машины.
Букса 5 крепится к цилиндру 4 кольцом 2 и стопорится винтом 3. Букса в цилиндре уплотнена резиновым кольцом, а шток с поршнем 8 уплотнен манжетами 9.
К цилиндру крепятся маслопроводы с гидрозамком 6 и дросселем 7, предназначенным для более плавной укладки бурильного оборудования при переводе из рабочего положения в транспортное.
Рис. 17. Замок гидравлический
Гидрозамок 6 служит для запирания жидкости в полостях гидроцилиндра при прекращении подачи рабочей жидкости.
Устройстве гидрозамка показано на рис. 17. В корпусе 1 размещены два обратных (запорных) шариковых клапана 4, между которыми расположен плавающий поршень 3 и толкатель 2. Рабочая жидкость под давлением поступает к замку через полости «Б» и «В» и от замка и гидроцилиндру через полости «А» и «Г».
Разборка гидрозамка категорически запрещена.
Рис. 18. Гидродомкрат
Гидродомкраты 7 (см. рис. 14) предназначены для разгрузки заднего моста автомобиля при бурении и установке опор. Каждый гидродомкрат имеет управление от автономной станции распределителя.
Гидродомкрат (рис. 18) состоит из штока 1, закрепленного неподвижно на раме машины: гильзы 3, совершающей возвратно- поступательное движение. Поршень 5 на штоке крепится гайкой 6. Направляющая штока 4 крепится упорным кольцом 8 и крышкой 2 с тремя болтами. Опора 7 шарнирно крепится к гильзе.
Рис. 19. Гидроцилиндр фрикциона
Гидроцилиндр (рис. 19) предназначен для периодического включения и выключения фрикциона. Цилиндр установлен на раздаточной коробке, шток цилиндра связан с рычагом отводки фрикционной муфты. К корпусу 3 цилиндра фрикциона крепятся крышки 2 и 4. Шток 1 с поршнем и крышки уплотнены резиновыми кольцами круглого сечения.
Штанга бурильная 13 (см. рис. 14) предназначена для осуществления возвратно-поступательного движения бурильного инструмента. Конструкция бурильной штанги представлена на рис. 8.
Рис. 20. Дроссель
Рис. 21. Линейный фильтр
Дроссель ПГ 77-14, предназначенный для регулирования скорости и усилия подачи бурильной штанги, установлен на пульте управления. Он состоит из корпуса 1 (рис. 20), втулки-дросселя 2, лимба 3, гайки фиксации лимба 4.
Расход рабочей жидкости регулируется изменением дросселирующей щели за счет перемещения втулки-дросселя 2 при повороте лимба. Лимб поворачивается на четыре оборота.
Линейный фильтр 1.1.32-2514 предназначен для очистки рабочей жидкости от механических примесей и установлен на сливной магистрали гидравлической системы.
Фильтр состоит из отстойника 1 (рис. 21), крышки 2 предохранительного клапана (запорного устройства) 3, втулки 4, резиновых колец 5, 11; фильтрующего элемента 6, опорной шайбы 7, пружины 8, пробки 9, индикатора загрязнений 10.
Индикатор загрязнения, установленный на фильтре, отрегулирован на давление 0,25+0,05 МПа (2.5+0,5 кгс/см2). При увеличении давления загорается сигнальная лампочка на пульте управления.
Предохранительный клапан предназначен для предотвращения выхода из строя фильтрующего элемента, а также для предотвращения вытекания рабочей жидкости из трубопроводов и масляного бака при замене фильтроэлементов.
Предохранительный клапан отрегулирован на давление 0,3±0,05 МПа (3±0,5 кгс/см2).
Регулировку и разборку индикатора загрязнения и предохранительного клапана производить запрещается.
| Групповые ссылки VIZITПартнеры группы VIZIT |
|
Эмбриональное происхождение и компартментальная организация наружных гениталий
Внешнее генитальное поле в задней латеральной пластинке мезодермы и переднем хвосте
В экспериментах по картированию судьбы, описанных выше, латеральные мезодермальные клетки между задней областью зачатка задней конечности и передней частью области хвоста дают начало генитальному бугорку, демонстрируя существование поля внешних половых клеток-предшественников (зоны 2 и 3 на рис.3). Примечательно, что клетки, помеченные на переднем крае зоны 2, но немного медиальнее края боковой пластинки, вносили вклад как в генитальный бугорок, так и в вентральную сторону зачатка задней конечности, предполагая, что внешнее генитальное поле расположено латеральнее и может частично перекрываются, с полем задних конечностей. Эти результаты в целом согласуются с результатами, полученными в параллельном исследовании Tschopp et al. 18 . Предыдущие исследования картирования судеб зачатков конечностей показали, что предшественники зачатков вентральных конечностей происходят из латеральной стороны латеральной пластинки, а предшественники зачатков дорсальных конечностей происходят из клеток в более медиальных положениях 15,16,17 .Наши результаты показывают, что регионализация вдоль медиолатеральной оси латеральной пластинки распространяется на третью популяцию клеток, внешние генитальные клетки-предшественники, которые расположены латеральнее вентральных предшественников задних конечностей (Fig. 6a). Во время закрытия стенки тела эти самые боковые клетки перемещаются к вентральной средней линии, где они дают начало парным генитальным набуханиям и, позже, генитальному бугорку (Fig. 6b).
Рисунок 6Регионализация латеральной мезодермы на дорсальную конечность, вентральную конечность и внешние генитальные поля вдоль медиолатеральной оси.
Схематическое изображение поперечных сечений клоакального уровня эмбриона до (а) и после (б) закрытия стенки тела. (а) Клетки-предшественники наружных половых органов (синие) берут начало на боковых краях латеральной мезодермы, прилегающих к вентральному полю конечностей (белые). Поле спинных конечностей показано красным. (b) Закрытие стенки тела объединяет левое и правое внешние генитальные поля на вентральной средней линии, где они дают начало парным генитальным набуханиям, которые образуют генитальный бугорок (синий).Слева и справа показаны зачатки конечностей, при этом брюшная конечность показана белым, а дорсальная конечность — красным.
Компартментальная организация генитального бугорка
Двойное мечение левой и правой латеральных пластинок мезодермы на уровне задних конечностей показало, что генитальный бугорок развивается из двух пулов предшественников, которые происходят по обе стороны от эмбриона. Когда стенка тела смыкается, левая и правая популяции латеральной пластинки мезодермы сближаются, сливаясь на вентральной средней линии.Парные генитальные набухания, которые появляются по обе стороны от клоакальной мембраны, происходят из левого и правого пулов генитальных предшественников, и они остаются разделенными клоакальной энтодермой, которая формирует эпителий уретральной пластинки. Неожиданно, после того, как парные генитальные опухоли сливаются, чтобы сформировать единый генитальный бугорок, клетки, происходящие с левой и правой сторон мезодермы латеральной пластинки, остаются ограниченными своими соответствующими сторонами происхождения, и небольшое перемешивание происходит по средней линии. Основываясь на этих наблюдениях, мы предполагаем, что генитальный бугорок демонстрирует истинную компартментальную организацию, sensu Drosophila компартментов, в которых соседние клеточные популяции обнаруживают клонное ограничение и соблюдают границу компартмента 19 (в данном случае эпителий уретральной пластинки).В сочетании с нашими недавними выводами о том, что происхождение генитального бугорка из парных опухолей половых органов является общей чертой амниот 4,5,6,7,8,9,10 , эти результаты повышают вероятность того, что организация левого и правого отделов генитального бугорка может быть широко консервативным признаком амниот наружных гениталий.
Регионализация латеральной мезодермы: расположение задних конечностей и гениталий
Парные гениталии амниот различаются по их переднезаднему и медиолатеральному положению (см.4). У птиц и млекопитающих, например, парные вздутия на гениталиях появляются кпереди от клоаки, тогда как у чешуек (ящериц и змей) парные вздутия возникают кзади и латеральнее клоаки 5,9 . Поразительный сдвиг наблюдается у зеленой ящерицы Anolis carolinensis , у которой начинается парное вздутие половых органов на вентрально-проксимальном конце зачатков ног 9 . В контексте представленных здесь результатов эта вариация предполагает, что граница между конечностью и полями гениталий в латеральной пластинке мезодермы может быть лабильной, и медиолатеральное смещение этой границы может объяснять вариации в положении половых органов относительно задних конечностей. и клоака.Наше открытие, что клетки, помеченные на медиальной стороне зоны 2, позже были обнаружены в генитальном бугорке и на вентральной стороне заднего зачатка задней конечности, предполагает, что задняя конечность и внешние генитальные поля могут частично перекрываться или иметь мягкую, а не резкую границу ( хотя не исключено, что эти инъекции метили клетки по обе стороны от границы). Независимо от того, примыкают ли поля внешних гениталий и задних конечностей или частично перекрываются, результаты, представленные здесь и в картах судьбы предшественников конечностей 15,16,17 демонстрируют, что пулы задних конечностей и внешних генитальных предков обычно являются отдельными популяциями клеток.Вариация относительного положения этих придатков может быть результатом сдвигов в положениях полей-предшественников, хотя мы не можем исключить возможность того, что поля конечностей и гениталий обычно фиксированы, и именно положения сигналов выроста относительно этих полей являются лабильными. .
У чешуйчатых парные гениталии остаются раздельными на протяжении всего развития, давая начало левым и правым гемипенам и гемиклиторам 4,5,9 . Мы не проследили происхождение плоских гемифаллусов; однако наши результаты предполагают, что каждый гемифалл может полностью возникнуть из клеток-предшественников на одной стороне эмбриона.Учитывая, что единичные срединные фаллосы млекопитающих 11 , архозавров 7,8,10 и черепах 6 развиваются в результате слияния левых и правых половых органов, наши результаты по клеточным линиям могут подтвердить идею, что чешуйчатые внешние половые структуры верны « hemi » (половинные) фаллосы. Эта интерпретация не предназначена для предположения структурной гомологии левого и правого гемипениса с левой и правой стороной срединных фаллосов у некваматных амниот, а скорее для того, чтобы подчеркнуть взаимосвязь между левым и правым полями гениталий в латеральной пластинке мезодермы и изменчивость морфологии фаллоса амниот.
Наше наблюдение, что клетки, меченные в зонах 2 и 3, вносят вклад в дорсальную (переднюю по отношению к туловищу) и вентральную (заднюю) стороны генитального бугорка, соответственно, предполагает, что внешние генитальные клетки-предшественники сохраняют свои исходные переднезадние положения. Эти паттерны клеточного поведения напоминают переднезаднюю организацию др. Структур, таких как лопатка куриной лопатки, в которой клетки сохраняют те же относительные положения, что и сомиты, из которых они происходят 20 .Основываясь на наших выводах о том, что клетки генитального бугорка происходят из самой задней боковой мезодермы туловища, смежной с задними клетками зачатка задних конечностей, есть соблазн предположить, что позиционная идентичность внешних генитальных предков, как и предшественников конечностей, определяется во время переднезадней регионализации ось основного корпуса. Действительно, связанные с AbdB гены Hox экспрессируются на заднем конце эмбриона, в генитальном бугорке и в задних областях зачатков конечностей, и возможно, что общая регуляция этих генов в конечностях и гениталиях 21 отражает их эмбриональное происхождение на одном осевом уровне туловища.
Компартментализация гениталий слева и справа и связь с комплексом эписпадия-экстрофия
Закрытие стенки тела во время эмбриогенеза имеет решающее значение для ограждения внутренних органов, интернализации кишечника и образования стебля желтка 22 . Этот процесс превращает эмбрион из плоского листа в трехмерную трубку (рис. 6). Застежка на стенке тела «застегивает» вместе левый и правый боковые края эмбриона по средней линии вентральной части. Этот процесс происходит как в переднем, так и в заднем направлении между анусом и глоткой 22 .Неполное закрытие стенки тела приводит к врожденным дефектам, таким как омфалоцеле, гастрошизис и комплекс экстрофии-эписпадии (EEC) 22,23 . Было высказано предположение, что этот процесс требует пролиферации и латеральной миграции мезодермального и эктодермального слоев латеральной пластинки, что заставляет складки тела расширяться и встречаться на вентральной средней линии 22,23,24,25,26,27 . Боковое складывание и круговая (от дорсальной к вентральной) миграция трех зародышевых листков (эктодермы, мезодермы и энтодермы) связаны с закрытием кишечника и мочеполовых синусов и образованием клоакальной мембраны сзади.
Наше открытие, что генитальный бугорок возникает из левого и правого половых предшественников, которые объединяются во время закрытия стенки тела, дает новое объяснение связи между эписпадией и экстрофией. Эписпадия — это эктопическое дорсальное отверстие уретры (в отличие от более распространенной гипоспадии, при которой уретра открывается эктопически на вентральной стороне полового члена) 28 . Эписпадия может проявляться как дорсальная щель полового члена или раздвоение клитора и часто связана с экстрофией мочевого пузыря, при которой мочевой пузырь выступает через вентральную стенку тела.Наше открытие, что фаллос имеет сложное происхождение, в котором левая и правая стороны представляют собой ограниченные по клону компартменты клеток-предшественников, которые объединяются во время закрытия стенки тела, предполагает, что общий механизм, включающий нарушение закрытия задней стенки тела, может лежать в основе обеих эписпадий. и экстрофия, и это может объяснить их частую ассоциацию у людей.
Границы | Последние достижения в области нейронных цепей для восприятия вкуса при голоде
Введение
Потребление пищи имеет важное значение для выживания, а нервные процессы, опосредующие информацию о голоде, направляют животных к поиску аппетитной пищи и последующему полноценному кормлению.Вкус важен для животных, чтобы оценить ценность пищи (Lindemann, 2001). Он играет несколько ролей как в аппетитном, так и в полноценном поведении. Многие области мозга демонстрируют разные вкусовые реакции при голоде и сытости, что указывает на то, что вкусовое восприятие модулируется внутренним состоянием. В этом обзоре мы кратко представим исследования вкусового восприятия в периферической и центральной нервной системе, а затем сосредоточимся на недавних открытиях о нервных цепях, связанных с модуляцией сладкого или горького вкуса при голоде, и о том, как они участвуют в кормлении.
Ощущение вкуса от языка к мозгу
Люди и многие млекопитающие способны распознать пять основных вкусов: сладкий, умами, горький, кислый и соленый. Вкусовые вещества обнаруживаются вкусовыми сосочками на языке. Это структуры в форме луковицы, состоящие из клеток, которые экспрессируют различные типы вкусовых рецепторов. Например, сладкие вещества обнаруживаются комбинацией рецепторов, связанных с G-белком T1R2 и T1R3, в то время как горькие вещества обнаруживаются рецепторами T2R (Zhao et al., 2003; Мейерхоф и др., 2005; Мюллер и др., 2005). Как показано на рисунке 1, информация о вкусе затем отправляется в ствол мозга через вкусовый ганглий в область, называемую ростральной частью ядра солитарного тракта (rNTS) (Beckstead and Norgren, 1979). У грызунов существует ретранслятор от rNTS к парабрахиальным ядрам (PBN), которые, в свою очередь, проецируют информацию на вентральное заднемедиальное таламическое ядро (VPMpc) (Beckstead et al., 1980; Pritchard et al., 1989). В конечном итоге информация о вкусе поступает в первичную вкусовую кору, которая также называется островковой корой (InsCtx), и во вторичную вкусовую кору, которая также называется орбитофронтальной корой (OFC) (Rolls et al., 1990). Хотя исследователи знали о нейронных путях вкуса на протяжении многих лет, молекулярная идентичность отдельных вкусовых нейронов в головном мозге была обнаружена редко. Некоторые недавние исследования выявили вкусовые нейроны, которые передают специфическую информацию о вкусе у мышей. Pdyn-экспрессирующие нейроны в rNTS реагируют исключительно на кислый вкус, и оптогенетическая активация этих нейронов вызывает отвращение (Zhang et al., 2019). В трехпортовом аппарате мышам был предоставлен доступ к среднему порту для слизывания различных растворов (воды, горького раствора или кислого раствора).Их обучали переходить влево (горький раствор или вода) или в правое отверстие (кислый раствор), чтобы сообщить об идентичности вкуса. В то время как мыши правильно сообщали, переходя к левому порту после слизывания воды или горького раствора, мыши, активированные Pdyn-нейронами, перешли к правому порту после слизывания воды, чтобы сообщить о кислом вкусе, предполагая, что эти нейроны необходимы для распознавания кислого вкуса. Другое исследование, посвященное области PBN, показало, что нейроны, экспрессирующие Satb2, избирательно реагируют на сладкий вкус и передают информацию в VPMpc, чтобы вызвать аппетитное облизывание (Fu et al., 2019а). Поскольку жир, помимо подсластителей, сильно стимулирует аппетит к еде, он повысил вероятность того, что вкус жира станет шестой модальностью вкуса. Хотя ранее считалось, что пероральное восприятие жирных кислот в основном зависит от текстуры и обоняния (Rolls et al., 1999), недавние исследования показали, что некоторые молекулы, такие как CD36 и GPR120, играют роль в обнаружении жирных кислот в периферических вкусовых системах. В CD36-положительных вкусовых клетках вкусовых рецепторов концентрация Ca 2+ увеличивалась после применения длинноцепочечных жирных кислот (Gaillard et al., 2008). Увеличение ответа Ca 2+ на линолевую кислоту наблюдалось во вкусовых клетках, экспрессирующих GPR120, у мышей (Ozdener et al., 2014). Более того, электрофизиологические записи вкусового нерва хорды барабанной перепонки мыши показывают, что GPR120 может играть роль в различении вкуса жирных кислот от других первичных вкусов (Yasumatsu et al., 2019).
Рисунок 1 . Нейронная регуляция вкусового восприятия в состоянии голода. Нейроны AgRP, которые сначала активируются в условиях голода, вызывают поиск пищи и, как следствие, потребительское поведение. (1) Нейроны AgRP ингибируют InsCtx через цепи AgRP-PVT-BLA-InsCtx для усиления упреждающей реакции на пищевой сигнал в процессе аппетитного кормления. (2) После начала еды вкус определяется языком и воспринимается вкусовой корой (InsCtx и OFC) через rNTS, PBN и VPMpc. (3) InsCtx кодирует положительную или отрицательную валентность вкуса, взаимодействуя с BLA или центральной миндалевидным телом, чтобы продолжить или прекратить кормление.OFC объединяет вкус и другую сенсорную информацию, а также ценность вознаграждения, проецируя ее на BLA. (4) Возбуждающие нейроны, экспрессирующие Vglut2 в латеральном гипоталамусе, также получают тормозящие сигналы от нейронов AgRP, чтобы вызвать питание в состоянии голода. Нейроны Vglut2 в LH проецируются на LS или на LHb, чтобы модулировать вкусовые или аверсивные вкусовые предпочтения соответственно. Вышеупомянутые области в значительной степени взаимодействуют с мезолимбической системой среднего мозга, создавая поощрительное обучение для регуляции пищевого поведения.BLA, базолатеральное миндалевидное тело, CeA: центральное ядро миндалины, InsCtx: кора островка; LHb — боковая габенула; LS — боковая перегородка; VPMpc, вентрально-заднемедиаль таламуса; OFC, орбитофронтальная кора; PBN, парабрахиальное ядро; rNTS, ростральная часть солитарного ядра.
Необходимо провести дальнейшие исследования, чтобы выяснить молекулярную идентичность других нейронов, связанных с путями вкусовых реакций, чтобы отобразить полную сеть ощущения вкуса в мозге.
Роль вкусовых ощущений в пищевом поведении
Вкусовые ощущения эволюционировали, чтобы служить доминирующим регулятором пищевого поведения (Ярмолинский и др., 2009). Поведение при приеме пищи можно разделить на гомеостатическое кормление для поддержания массы тела и метаболических функций или гедоническое кормление, основанное на сенсорном восприятии или удовольствии (Berthoud, 2004; Rossi and Stuber, 2018). Примечательно, что гомеостатическая и гедонистическая системы кормления активируются отдельно или одновременно в зависимости от различных ситуаций, связанных с кормлением (Castro et al., 2015). Паттерн активации этих двух систем может меняться в зависимости от вкуса пищи (приятный или неприятный), а также в соответствии с физиологическим состоянием животного (голод или сытость). Животные начинают пищевое поведение в состоянии голода и прекращают кормление, когда их кормят до сытости. Этот процесс регулируется гомеостатической системой, чтобы предотвратить избыточное потребление калорий (Campos et al., 2016). Однако вкусная еда может активировать мезолимбическую систему вознаграждения, чтобы стимулировать прием пищи даже в сытом состоянии.Гедоническое кормление вызывает аппетитное поведение, такое как тяга к сахару, что приводит к перееданию (Hajnal et al., 2004).
Вкусовые ощущения и внутреннее состояние
Голодные и сытые животные по-разному реагируют на вещества с привлекательным или потенциально вредным вкусом. Нейровизуализационные исследования на людях показали, что в состоянии голода области мозга, связанные с системой вознаграждения, были более активными после представления очень вкусной пищи по сравнению с теми же областями во время насыщения.Это говорит о том, что пища становится более вкусной при голоде, что описывается старой поговоркой «Голод — лучшая специя» (Siep et al., 2009).
Предыдущее исследование, включающее задачу оценки вкуса человека, показало, что пороги распознавания сахарозы и соли были значительно ниже во время голодания, чем во время насыщения (Зверев, 2004). Другое исследование показало, что участники проявляли значительно более высокую чувствительность к сладкому, кислому и соленому вкусу во время состояния голода и более высокую чувствительность к горькому вкусу во время насыщения (Hanci and Altun, 2016).Эти результаты показывают, что внутреннее состояние может напрямую влиять на сенсорное восприятие вкуса.
Некоторые исследования также показали, что вкусовые качества или стимулирующая ценность вкуса зависят от чувства голода. Голодные люди демонстрируют повышенную самооценку субъективных чувств по отношению к приятному вкусу (Rolls et al., 1983). Другое исследование показало, что у крыс после голодания увеличилось пристрастие к сладкому вкусу (Berridge, 1991). Авторы оценивали приятность вкуса, используя мимику вкусовой реакции.Крысы, лишенные пищи на 48 ч, показали более высокие показатели вкусовой реактивности для смеси сахароза-хинин.
В нескольких исследованиях сообщалось, что гормоны и нейропептиды, связанные с кормлением, могут модулировать периферическую вкусовую чувствительность. Сообщается, что среди различных гормонов анорексии лептин избирательно ингибирует ответы, связанные со сладким вкусом, в клетках вкусовых рецепторов, связываясь с функциональным рецептором лептина Ob-Rb в грибовидных и окаймляющих вкусовых рецепторах (Kawai et al., 2000; Shigemura et al., 2004; Йошида и др., 2015). Глюкагоноподобный пептид 1 (GLP-1) увеличивает чувствительность к сладкому вкусу через рецепторы GLP-1, расположенные на афферентных нервных волокнах, прилегающих к вкусовым сосочкам (Shin et al., 2008), в то время как холецистокинин (CCK) на горький вкус через CCK -A рецепторы экспрессируются в клетках вкусовых рецепторов (Herness et al., 2002; Lu et al., 2003). Более того, инсулин, по-видимому, усиливает реакцию на соленый вкус через эпителиальные натриевые каналы (ENaC) (Baquero and Gilbertson, 2011). Интраназальное введение инсулина увеличивало чувствительность к сладкому, горькому, соленому, кислому вкусу в сенсорных тестах человека (Rodriguez-Raecke et al., 2017). Нейропептид Y, известный как орексигенный пептид, может влиять на чувствительность к горькому вкусу через NPY1R в клетках вкусовых рецепторов (Zhao et al., 2005). Сообщается, что окситоцин снижает чувствительность к сладкому у мышей через рецепторы окситоцина, расположенные во вкусовых сосочках (Sinclair et al., 2015). Однако центральный механизм модуляции вкуса при голоде до сих пор неясен.
Области мозга, демонстрирующие вкусовую реакцию, зависящую от голода
В прошлом исследования функциональной магнитно-резонансной томографии (фМРТ) или электрофизиологии открывали области мозга, включая OFC, миндалевидное тело, InsCtx и боковой гипоталамус (LH), которые реагируют на вкусовые сигналы в зависимости от голода / насыщения.Исследование на людях с помощью нейровизуализации показало, что насыщенные участники сообщали об ослабленной субъективной приятности определенного вкуса, предполагая, что сытость отрицательно влияет на восприятие вкуса (Rolls et al., 1983; Kringelbach et al., 2003). Это снижение приятного вкуса показало высокую корреляцию со снижением активности OFC.
Электрофизиологические исследования, проведенные на приматах, выявили большую популяцию нейронов в OFC (Nakano et al., 1984; Yamamoto et al., 1984; Rolls et al., 1989), миндалевидное тело (Nakano et al., 1986) и LH (Burton et al., 1976), которые реагировали на раствор сахарозы, когда животное было голодно, но не когда оно было насыщено. Важно отметить, что одновременная электрофизиологическая регистрация одиночных нейронов из OFC, LH и миндалевидного тела проводилась у крыс, лишенных пищи и имеющих свободный доступ к раствору сахарозы (de Araujo et al., 2006). Подобно результатам, полученным на приматах, крысы проявляли голодную реакцию на вкусы в этих областях мозга.Однако из-за ограничения разрешения изображений в исследованиях фМРТ или электрофизиологии точная роль этих областей мозга, которые показали зависящую от состояния вкусовую реакцию, остается в значительной степени неуловимой.
Нейронные цепи для восприятия вкуса и коррелированного пищевого поведения во время голода
В условиях голодания животным необходимо искать пищу, принимать решение о том, есть ли она или отказаться от нее, смакуя вкус, и потреблять пищу для выживания. Предполагается, что отдельные нейронные цепи, связанные с аппетитом, вкусовым восприятием и вознаграждением, работают вместе, вызывая аппетитное поведение и регулируя последующее консумативное поведение (Rolls, 2005; Ferrario et al., 2016).
Чтобы понять роль вкусовой системы во время состояния голода, пищевое поведение можно разделить на три фазы: (1) мотивационный и упреждающий процесс перед началом кормления (аппетитное поведение), (2) восприятие вкуса и выполнение кормления ( консумативное поведение) и (3) прекращение кормления (рис. 2). В последние годы недавно разработанные методы нейронной манипуляции и визуализация in vivo кальция позволили изучить нейронные функции, связанные с системой вкуса и регуляцией питания, с использованием высокого разрешения по времени.
Рисунок 2 . Роль внутреннего состояния и вкусовых качеств в кормлении. Голод побуждает животных инициировать поиск пищи и способствовать прогнозированию вкусовых качеств и ценности вознаграждения в процессе аппетитного кормления. В то время как приятный вкус приводит к потреблению пищи, неприятный вкус способствует ее отторжению. Этот процесс также способствует положительному или отрицательному подкреплению к пище. Голод усиливает предпочтение приятного вкуса, уменьшая при этом отвращение к неприятному вкусу, чтобы способствовать полноценному кормлению, а также препятствует прекращению кормления до насыщения.
Процесс мотивации и ожидания перед началом кормления
Голод стимулирует аппетитное поведение к поиску пищи и вызывает пищевое поведение (Sternson et al., 2013). Гипоталамус считался центром питания. Он содержит множество типов нейрональных клеток, связанных с поддержанием энергетического гомеостаза. Среди них нейроны, которые специфически экспрессируют связанный с Агути белок (AgRP) в дугообразном ядре гипоталамуса, были идентифицированы как голодные нейроны.Удаление нейронов AgRP у взрослых мышей приводит к афагии (Wu et al., 2009). И наоборот, использование оптогенетических и хемогенетических методов для быстрой активации нейронов, специфичных для клеточного типа, показало, что нейронов AgRP достаточно, чтобы вызвать пищевое поведение у кормленных мышей в течение нескольких минут, а количество потребляемой пищи аналогично тому, которое наблюдается у голодных мышей в течение ночи (Krashes et al., 2011; Betley et al., 2013). Интересно, что визуализация кальция нейронов AgRP на основе GCaMP с использованием волоконной фотометрии показала, что активность этих нейронов начинает снижаться еще до того, как вы перекусили (Chen et al., 2015), предполагая, что нейроны AgRP участвуют в упреждающем процессе для прогнозирования потребления калорий. Эта упреждающая реакция на пищевые сенсорные сигналы может помочь процессу перехода от поиска пищи к последующему кормлению.
Исследования фМРТ человека показали, что реакция InsCtx или OFC на сладкие растворы изменяется в зависимости от голода (Haase et al., 2009). Голодным или сытым участникам давали орально вкусовые растворы. Зависимое от уровня оксигенации крови изменение сигнала, измеренное с помощью фМРТ, показало значительные различия в активации сахарозы, кофеина, сахарина и лимонной кислоты в OFC и в InsCtx между голодом и насыщением.Недавние исследования (Livneh et al., 2017, 2020) продемонстрировали, что определенный путь от стимулирующих голод нейронов AgRP к InsCtx через паравентрикулярный таламус (PVT) и базолатеральную миндалину (BLA) участвует в зависимом от голода усилении пищевые подсказки у мышей (рис. 1-1). Авторы применили задачу визуального различения, в которой мышей обучали лизать после различных выученных визуальных сигналов, связанных с доставкой сладкого раствора (Ensure), горького раствора (хинин) или воды.Визуализация одноклеточного кальция нейронов InsCtx на основе GCaMP у ведущих мышей показала, что большая популяция нейронов InsCtx реагировала на визуальный сигнал пищи, лизание или сладкий вкус. Авторы использовали хинин для тренировки задачи визуального различения, чтобы позволить мышам усвоить отрицательный пищевой сигнал. Однако во время реальной визуализации мышам доставляли только Ensure. Интересно, что большая часть нейронов, связанных с ответом на пищевые сигналы, сильно активировалась, когда мыши слизывали сладкий раствор.Визуализация одних и тех же нейронов InsCtx во время голода и насыщения показала, что нейроны, которые реагировали на визуальные сигналы во время голода, исчезали во время насыщения. Другое исследование человека с помощью фМРТ показало, что просмотр изображений вкусной еды может активировать InsCtx (Simmons et al., 2005). Эти данные свидетельствуют о том, что InsCtx представляет собой прогноз питания и интероцептивные последствия предстоящего потребления. Таким образом, эта гипоталамус-корковая связь показывает возможность улучшения вкусовых качеств пищи во время состояния голода.
Исследование человека с помощью фМРТ показало, что OFC и миндалевидное тело кодируют прогнозируемую ценность вознаграждения (Gottfried et al., 2003). In vivo исследование кальциевой визуализации нейронов OFC (Jennings et al., 2019), которые являются прямыми проецируемыми мишенями InsCtx, продемонстрировало значительные возбуждающие реакции во время облизывания калорийным вознаграждением у голодных мышей. Сочетание каждой доставки калорийного вознаграждения с оптогенетической стимуляцией чувствительных к кормлению клеток значительно увеличивало вылизывание. Интересно, что стимуляция чувствительных к питанию клеток не увеличивает вылизывание некалорийного подсластителя сахарина, что позволяет предположить, что нейроны OFC реагируют на содержание калорийной награды, а не на вкусовую награду.Также сообщается (Malvaez et al., 2019), что нейроны OFC, проектирующие BLA, кодируют зависящую от состояния стимулирующую ценность приятного пищевого вознаграждения. Активность рецептора глутамата в BLA необходима для кодирования и извлечения значения вознаграждения. Проекции от бокового OFC к BLA необходимы и достаточны для кодирования положительного значения вознаграждения. С другой стороны, проекции от медиального OFC к BLA необходимы и достаточны для извлечения этого значения из памяти. Исследование парадигмы поощрения девальвации показало, что OFC и BLA играют разные роли в обеспечении нормальной целенаправленной деятельности (Pickens et al., 2003). BLA кажется критически важным для формирования представлений, которые связывают реплики со стимулирующими свойствами результатов, но OFC, возможно, способствует сохранению этих представлений в памяти и обновлению их новой информацией.
Эти результаты означают, что InsCtx и OFC не только реагируют на вкусовые особенности, но также играют роль в зависимом от состояния предсказании вознаграждения перед кормлением. В то время как InsCtx и OFC, возможно, участвуют в прогнозировании вкуса, а также пищевого вознаграждения, OFC содержит определенную популяцию нейронов, которая способствует прогнозированию калорийности.
Восприятие вкуса во время кормления
Пищевые сигналы, такие как визуальная информация и запах, позволяют животным предсказывать вкусовые качества, а также вознаграждение за еду (Rolls, 2015). Решение о фактическом оральном кормлении зависит от вкуса. Вкусовые пути от ствола мозга к кортикальным областям важны для создания ощущения вкуса (рис. 1-2). Оральный контакт с пищей с нормальным вкусом поддерживает кормление, в то время как приятный вкус вызывает положительные ощущения и увеличивает частоту жевания.Однако неприятные вкусы передают негативные ощущения и приводят к резкому прекращению кормления.
Недавнее исследование (Wang et al., 2018) продемонстрировало, что иннервация между InsCtx и миндалевидным телом помогает передавать вкусовые валентности (рис. 1-3). Вкусовые коры сладкого и горького вкусов расположены в передней и задней части InsCtx. Они иннервируют отдельные субъядра в миндалевидном теле (BLA для сладкого и центральное ядро миндалины [CeA] для горького).Оптогенетическая активация нейронов InsCtx, проецирующих BLA, усиливает поведение при облизывании воды, вызывая «виртуальное» ощущение сладкого вкуса, в то время как активация нейронов InsCtx, проецирующих CeA, уменьшает слизывание по направлению к воде, имитируя горький вкус. Другое исследование с использованием теста «годен / не годен», усиленного вкусовыми добавками (сахароза / хинин), показало, что специфическое ингибирование высвобождения нейромедиаторов из латеральных нейронов InsCtx, проецирующих CeA (CeL), не позволяло мышам приобретать ответ «нет» на хинин и нарушил ответную реакцию на сахарозу в задаче «годен / не годен», что указывает на то, что цепь InsCtx-CeL важна для установления поведенческой реакции на сигналы, предсказывающие аппетитные или аверсивные вкусовые вещества (Schiff et al., 2018). Эти результаты предполагают, что InsCtx содержит нейроны, которые кодируют врожденную валентность вкуса, а также выученное предсказание вкусовых подсказок, чтобы вызвать аппетит или отторжение. Однако неясно, отличаются ли нейроны, связанные с вкусовым ответом в InsCtx, от нейронов, предсказывающих сигналы в InsCtx, или перекрываются с ними.
Принимая во внимание доказательства связи между миндалевидным телом и мезолимбической системой вознаграждения, миндалевидное тело может функционировать как ретрансляционная точка, передающая валентность вкуса и свойство вознаграждения (InsCtx-миндалевидное тело и OFC-миндалевидное тело) для запуска пищевого поведения.
Модуляция вкуса и внутреннего состояния для прекращения кормления
Во время дефицита энергии животным необходимы калории для поддержания веса тела и метаболических функций. Они продолжают полноценное питание от состояния голода до насыщения, чтобы удовлетворить потребность в калориях. ЛГ является важной нейроанатомической областью как для аппетитного, так и для потребительского поведения (Nieh et al., 2015). ЛГ содержит множество различных типов нейронов, а активация ГАМКергических (экспрессирующих Vgat) нейронов в ЛГ усиливает как аппетитное, так и консумативное поведение.Визуализация кальция у свободно ведущих мышей показала, что отдельные нейроны Vgat LH преимущественно кодируют аспекты аппетитного или потребного поведения (Jennings et al., 2015). Напротив, глутаматергические нейроны Vglut2, которых также много в ЛГ, играют противоположную роль в регулировании пищевого поведения. Оптогенетическая стимуляция нейронов Vglut2 LH не только снижает аппетит у голодных мышей, но также вызывает отвращение к участкам, связанным со стимуляцией этих клеток (Stamatakis et al., 2016). In vivo исследование с визуализацией кальция на нейронах Vglut2 LH продемонстрировало больший ответ во время доставки сахарозы при насыщении по сравнению с ответом в состоянии голода (Rossi et al., 2019). Хотя эти результаты противоположны результатам ФМРТ или электрофизиологических исследований ЛГ, показывающих большую реакцию на голод, чем на сытость, можно предположить, что в нейронах LH и Vglut2 существуют разные популяции нейронов, которые специфически кодируют чувство насыщения и служат тормозом для прекращения кормление действие.
Электрофизиологическое исследование (Li et al., 2013) показало, что ЛГ содержит две отдельные популяции нейронов, отвечающих на аппетитные и аверсивные вкусы. Недавнее исследование показало, что физиологический голод влияет на предпочтения аппетитных и отталкивающих вкусов в мышиной модели. Эти эффекты индуцируются LH-проектирующими нейронами AgRP (Fu et al., 2019b). В краткосрочном вкусовом тесте хемогенетическая и оптогенетическая активация нейронов AgRP у накормленных животных увеличивала количество лизаний для раствора, содержащего относительно низкую концентрацию сахарозы.Напротив, увеличение количества лизаний для горького раствора (денатония) также наблюдалось после оптогенетической активации нейронов AgRP. Кроме того, хемогенетическое ингибирование нейронов Vglut2 LH , но не нейронов Vgat LH , повторяет индуцированную голодом модификацию вкуса. Два различных нейронных пути от нейронов Vglut2 LH к латеральной перегородке или к латеральной габенуле (LHb) вносят вклад в модуляцию аппетитного и аверсивного вкуса, соответственно.LS и LHb являются частями системы мезолимбического вознаграждения, что указывает на то, что LH может модулировать ценность вкуса в качестве вознаграждения (рис. 1-4). В то время как нейроны Vglut2 LH , которые кодируют чувство насыщения, служат тормозом для прекращения консумматического поведения во время состояния голода (Stamatakis et al., 2016; Rossi et al., 2019), эти нейроны получают тормозные сигналы от нейронов AgRP. Повышение предпочтения сладких веществ и толерантности к веществам с горьким вкусом за счет AgRP-зависимого ингибирования нейронов Vglut2 LH может способствовать консумативному поведению для удовлетворения возникающей потребности в калориях.
Примечательно, что OFC прямо и косвенно связан с LH (Öngür and Price, 2000; Reppucci and Petrovich, 2016), предполагая, что OFC может передавать интегрированную сенсорную информацию в LH, вызывая мотивацию к приятному вкусу. Кроме того, ЛГ взаимодействует с мезолимбической системой, чтобы стимулировать обучение за положительное или отрицательное подкрепление и модулировать вкусовые предпочтения, чтобы вызвать или предотвратить потребление пищи в будущем.
Заключение
Кормление регулируется гомеостатической и гедонической системами кормления.Вкусовые ощущения от ствола мозга до вкусовой коры служат регулятором и движущим фактором питания, взаимодействуя с гипоталамусом и мезолимбической системой вознаграждения. Мозг объединяет сенсорную информацию и интероцепцию, чтобы управлять правильным кормлением для выживания. Понимание механизма, с помощью которого мозг воспринимает вкус и способы вознаграждения в различных физиологических состояниях, может быть полезным в терапевтическом лечении переедания в будущем.
Авторские взносы
Все перечисленные авторы внесли существенный, прямой и интеллектуальный вклад в работу и одобрили ее к публикации.
Финансирование
ПроектKN был частично поддержан грантами на научные исследования (18H02160) Министерства образования, культуры, спорта, науки и Научным фондом Такеда.
Конфликт интересов
Авторы заявляют, что исследование проводилось при отсутствии каких-либо коммерческих или финансовых отношений, которые могут быть истолкованы как потенциальный конфликт интересов.
Список литературы
Бакеро, А. Ф., и Гилбертсон, Т.А. (2011). Инсулин активирует эпителиальный натриевый канал (ENaC) через фосфоинозитид-3-киназу в клетках вкусовых рецепторов млекопитающих. г. J. Physiol. Cell Physiol. 300, C860 – C871. DOI: 10.1152 / ajpcell.00318.2010
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Бекстед Р. М., Морс Дж. Р. и Норгрен Р. (1980). Ядро одиночного тракта у обезьяны: выступы к таламусу и ядра ствола мозга. J. Comp. Neurol. 190, 259–282.DOI: 10.1002 / cne.9015
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Бекстед Р. М. и Норгрен Р. (1979). Авторадиографическое исследование центрального распределения тройничного, лицевого, языкоглоточного и блуждающего нервов у обезьяны. J. Comp. Neurol. 184, 455–472. DOI: 10.1002 / cne.
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Бетли, Дж. Н., Цао, З. Ф. Х., Ритола, К. Д., Стернсон, С.М. (2013). Организация параллельных цепей с резервированием для гомеостатического контроля поведения при кормлении. Ячейка 155, 1337–1350. DOI: 10.1016 / j.cell.2013.11.002
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Бертон, М. Дж., Роллс, Э. Т. и Мора, Ф. (1976). Влияние голода на реакцию нейронов бокового гипоталамуса на вид и вкус пищи. Exp. Neurol. 51, 668–677. DOI: 10.1016 / 0014-4886 (76) -8
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Кампос, К.А., Боуэн, А. Дж., Шварц, М. В., и Палмитер, Р. Д. (2016). Парабрахиальные нейроны CGRP контролируют прекращение приема пищи. Cell Metab. 23, 811–820. DOI: 10.1016 / j.cmet.2016.04.006
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Кастро, Д. К., Коул, С. Л., и Берридж, К. С. (2015). Роль латерального гипоталамуса, прилежащего ядра и брюшной паллидума в еде и голоде: взаимодействие между гомеостатической схемой и схемой вознаграждения. Фронт. Syst. Neurosci. 9:90. DOI: 10.3389 / fnsys.2015.00090
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Чен, Ю., Лин, Ю. К., Куо, Т. В., и Найт, З. А. (2015). Сенсорное обнаружение пищи быстро изменяет дугообразные схемы кормления. Мобильный 160, 829–841. DOI: 10.1016 / j.cell.2015.01.033
PubMed Аннотация | CrossRef Полный текст | Google Scholar
de Araujo, I.E., Gutierrez, R., Oliveira-Maia, A.J., Pereira, A., Nicolelis, M.A.L., и Simon, S.А. (2006). Нейронно-ансамблевое кодирование состояний сытости. Neuron 51, 483–494. DOI: 10.1016 / j.neuron.2006.07.009
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Феррарио, К. Р., Лабуэб, Г., Лю, С., Ние, Э. Х., Раус, В. Х., Сюй, С. и др. (2016). Гомеостаз отвечает мотивации в борьбе за контроль над приемом пищи. J. Neurosci. 36, 11469–11481. DOI: 10.1523 / JNEUROSCI.2338-16.2016
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Фу, О., Иваи, Ю., Кондо, К., Мисака, Т., Минокоши, Ю., Накадзима, К., и Итиро (2019a). Нейроны, экспрессирующие SatB2 в парабрахиальном ядре, кодируют сладкий вкус. Cell Rep. 27, 1650–1656.e4. DOI: 10.1016 / j.celrep.2019.04.040
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Fu, O., Iwai, Y., Narukawa, M., Ishikawa, A. W., Ishii, K. K., Murata, K., et al. (2019b). Гипоталамические нейронные цепи, регулирующие изменение вкуса, вызванное голодом. Nat. Commun. 10: 4560. DOI: 10.1038 / s41467-019-12478-x
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Gaillard, D., Laugerette, F., Darcel, N., El-Yassimi, A., Passilly-Degrace, P., Hichami, A., et al. (2008). Вкусовой путь участвует в CD36-опосредованном оросенсорном восприятии длинноцепочечных жирных кислот у мышей. FASEB J. 22, 1458–1468. DOI: 10.1096 / fj.07-8415com
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Готфрид, Дж.А., О’Догерти, Дж., И Долан, Р. Дж. (2003). Кодирование прогностической ценности вознаграждения в миндалевидном теле и орбитофронтальной коре человека. Наука 301, 1104–1107. DOI: 10.1126 / science.1087919
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Haase, L., Cerf-Ducastel, B., and Murphy, C. (2009). Активация коры головного мозга в ответ на стимулы чистого вкуса во время физиологических состояний голода и сытости. Neuroimage 44, 1008–1021. DOI: 10.1016 / j.neuroimage.2008.09.044
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Хайнал А., Смит Г. П. и Норгрен Р. (2004). Пероральная стимуляция сахарозой увеличивает уровень дофамина прилежащего вещества у крыс. г. J. Physiol. — Регул. Интегр. Комп. Physiol. 281, R31 – R37. DOI: 10.1152 / ajpregu.00282.2003
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Hanci, D., и Altun, H. (2016). Состояние голода влияет как на обонятельные способности, так и на вкусовую чувствительность. Eur. Arch. Ото-Рино-Ларингол . 273, 1637–1641. DOI: 10.1007 / s00405-015-3589-6
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Хернесс, С., Чжао, Ф. Л., Лу, С. Г., Кая, Н., и Шен, Т. (2002). Экспрессия и физиологические действия холецистокинина в клетках вкусовых рецепторов крыс. J. Neurosci. 22, 10018–10029. DOI: 10.1523 / JNEUROSCI.22-22-10018.2002
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Дженнингс, Дж.Х., Ким, К. К., Маршель, Дж. Х., Раффи, М., Йе, Л., Квирин, С. и др. (2019). Взаимодействующие нейронные ансамбли в орбитофронтальной коре головного мозга для социального и пищевого поведения. Природа 565, 645–649. DOI: 10.1038 / s41586-018-0866-8
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Дженнингс, Дж. Х., Унг, Р. Л., Ресендез, С. Л., Стаматакис, А. М., Тейлор, Дж. Г., Хуанг, Дж. И др. (2015). Визуализация динамики гипоталамической сети для определения аппетитного и потребного поведения. Ячейка 160, 516–527. DOI: 10.1016 / j.cell.2014.12.026
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Кавай К., Сугимото К., Накашима К., Миура Х. и Ниномия Ю. (2000). Лептин как модулятор чувствительности к сладкому вкусу у мышей. Proc. Natl. Акад. Sci. USA 97, 11044–11049. DOI: 10.1073 / pnas.1
697
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Крашес, М. Дж., Кода, С., Е, К. П., Роган, С.К., Адамс, А. С., Кушер, Д. С. и др. (2011). Быстрая обратимая активация нейронов AgRP определяет пищевое поведение мышей. J. Clin. Вкладывать деньги. 121, 1424–1428. DOI: 10.1172 / JCI46229
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Kringelbach, M. L., O’Doherty, J., Rolls, E. T., and Andrews, C. (2003). Активация орбитофронтальной коры человека на раздражитель жидкой пищи коррелирует с его субъективной приятностью. Cereb. Cortex 13, 1064–1071.DOI: 10.1093 / cercor / 13.10.1064
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Ли, Дж. Х., Йошида, Т., Монк, К. Дж., И Кац, Д. Б. (2013). Боковой гипоталамус содержит два типа вкусовых реакций, связанных с вкусовыми качествами, с четкой динамикой. J. Neurosci. 33, 9462–9473. DOI: 10.1523 / JNEUROSCI.3935-12.2013
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Ливне Ю., Рамеш Р. Н., Берджесс К. Р., Левандовски К.M., Madara, J.C., Fenselau, H., et al. (2017). Гомеостатические цепи избирательно контролируют пищевые сигналы коры островка. Природа 546, 611–616. DOI: 10.1038 / природа22375
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Livneh, Y., Sugden, A.U., Madara, J.C., Essner, R.A., Flores, V.I., Sugden, L.A., et al. (2020). Оценка текущего и будущего физиологического состояния островковой коры. Нейрон 105, 1094–1111.e10. DOI: 10.1016 / j.нейрон.2019.12.027
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Лу, С. Г., Чжао, Ф. Л., и Хернесс, С. (2003). Физиологическое фенотипирование холецистокинин-чувствительных клеток вкусовых рецепторов крыс. Neurosci. Lett. 351, 157–160. DOI: 10.1016 / j.neulet.2003.07.016
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Мальваез М., Ши К., Мерфи М. Д., Гринфилд В. Ю. и Вассум К. М. (2019). Четкие проекции коры головного мозга и миндалины управляют кодированием и извлечением значения вознаграждения. Nat. Neurosci. 22, 762–769. DOI: 10.1038 / s41593-019-0374-7
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Мюллер, К. Л., Хун, М. А., Эрленбах, И., Чандрашекар, Дж., Цукер, К. С., и Рыба, Н. Дж. П. (2005). Рецепторы и логика кодирования горечи. Природа 434, 225–229. DOI: 10.1038 / nature03352
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Накано, Ю., Оомура, Ю., Ленард, Л., Нишино, Х., Aou, S., Yamamoto, T., et al. (1986). Связанная с кормлением активность глюкозо- и морфин-чувствительных нейронов миндалины обезьян. Brain Res. 399, 167–172. DOI: 10.1016 / 0006-8993 (86)
-XPubMed Аннотация | CrossRef Полный текст | Google Scholar
Накано, Ю., Оомура, Ю., Нишино, Х., Ау, С., Ямамото, Т., и Немото, С. (1984). Активность нейронов в медиальной орбитофронтальной коре головного мозга обезьяны: модуляция глюкозой и насыщением. Brain Res.Бык. 12, 381–385. DOI: 10.1016 / 0361-9230 (84)
-6
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Ние, Э. Х., Мэтьюз, Г. А., Оллсоп, С. А., Пресбрей, К. Н., Леппла, К. А., Вичманн, Р. и др. (2015). Расшифровка нейронных цепей, контролирующих компульсивный поиск сахарозы. Мобильный 160, 528–541. DOI: 10.1016 / j.cell.2015.01.003
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Онгюр, Д., и Прайс, Дж. Л. (2000).Организация сетей внутри орбитальной и медиальной префронтальной коры крыс, обезьян и людей. Cereb. Cortex 10, 206–219. DOI: 10.1093 / cercor / 10.3.206
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Ozdener, M.H., Subramaniam, S., Sundaresan, S., Sery, O., Hashimoto, T., Asakawa, Y., et al. (2014). CD36- и GPR120-опосредованная передача сигналов Ca2 + в клетках вкусовых луковиц человека опосредует дифференциальные ответы на жирные кислоты и изменяется у мышей с ожирением. Гастроэнтерология 146, 995–1005. DOI: 10.1053 / j.gastro.2014.01.006
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Пикенс, К. Л., Саддорис, М. П., Сетлоу, Б., Галлахер, М., Холланд, П. К. и Шенбаум, Г. (2003). Различные роли орбитофронтальной коры и базолатеральной миндалины в задаче обесценивания подкрепления. J. Neurosci. 23, 11078–11084. DOI: 10.1523 / JNEUROSCI.23-35-11078.2003
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Причард, Т.К., Гамильтон Р. Б. и Норгрен Р. (1989). Нейронное кодирование вкусовой информации в таламусе Macaca mulatta. J. Neurophysiol. 61, 1–14. DOI: 10.1152 / jn.1989.61.1.1
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Репуччи, К. Дж., И Петрович, Г. Д. (2016). Организация связей между миндалевидным телом, медиальной префронтальной корой и латеральным гипоталамусом: исследование однократного и двойного ретроградного отслеживания на крысах. Brain Struct. Функц. 221, 2937–2962. DOI: 10.1007 / s00429-015-1081-0
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Rolls, E. T., Critchley, H. D., Browning, A. S., Hernadi, I., and Lenard, L. (1999). Ответы на сенсорные свойства жира нейронов орбитофронтальной коры приматов. J. Neurosci. 19, 1532–1540. DOI: 10.1523 / JNEUROSCI.19-04-01532.1999
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Роллс, Э.Т., Роллс, Б.Дж. И Роу Э. А. (1983). Сытность, специфичная для органов чувств и мотивации, для восприятия и вкуса пищи и воды у человека. Physiol. Behav. 30, 185–192. DOI: 10.1016 / 0031-9384 (83)
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Rolls, E. T., Sienkiewicz, Z. J., and Yaxley, S. (1989). Голод модулирует ответы на вкусовые стимулы отдельных нейронов каудолатеральной орбитофронтальной коры макака. Eur. J. Neurosci. 1, 53–60. DOI: 10.1111 / j.1460-9568.1989.tb00774.x
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Rolls, E. T., Yaxley, S., and Sienkiewicz, Z. J. (1990). Вкусовые ответы одиночных нейронов каудолатеральной орбитофронтальной коры макака. J. Neurophysiol. 64, 1055–1066. DOI: 10.1152 / jn.1990.64.4.1055
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Росси, М.А., Басири, М.Л., МакГенри, Дж.A., Kosyk, O., Otis, J.M., Van Den Munkhof, H.E., et al. (2019). Ожирение изменяет активность и состояние транскрипции латерального тормоза гипоталамуса при кормлении. Наука 364, 1271–1274. DOI: 10.1126 / science.aax1184
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Schiff, H.C., Bouhuis, A. L., Yu, K., Penzo, M. A., Li, H., He, M., et al. (2018). Цепь островка-центральная миндалины для управления предпочтительным поведением, усиленным вкусовыми добавками. J. Neurosci. 38, 1418–1429. DOI: 10.1523 / JNEUROSCI.1773-17.2017
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Шигемура, Н., Охта, Р., Кусакабэ, Ю., Миура, Х., Хино, А., Кояно, К. и др. (2004). Лептин модулирует поведенческие реакции на сладкие вещества, влияя на периферические вкусовые структуры. Эндокринология 145, 839–847. DOI: 10.1210 / en.2003-0602
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Шин, Ю.К., Мартин, Б., Голден, Э., Дотсон, К. Д., Модсли, С., Ким, В. и др. (2008). Модуляция вкусовой чувствительности с помощью передачи сигналов GLP-1. J. Neurochem. 106, 455–463. DOI: 10.1111 / j.1471-4159.2008.05397.x
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Сип, Н., Рофс, А., Робрук, А., Хаверманс, Р., Бонте, М. Л., и Янсен, А. (2009). Голод — лучшая специя: ФМРТ-исследование влияния внимания, голода и калорийности на переработку пищевого вознаграждения в миндалевидном теле и орбитофронтальной коре головного мозга. Behav. Brain Res. 198, 149–158. DOI: 10.1016 / j.bbr.2008.10.035
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Симмонс В. К., Мартин А. и Барсалу Л. В. (2005). Изображения аппетитных блюд активируют вкусовые ощущения коры головного мозга для вкуса и вознаграждения. Cereb. Cortex . 15, 1602–1608. DOI: 10.1093 / cercor / bhi038
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Синклер, М.С., Переа-Мартинес, И., Абуяред, М., Сент-Джон, С. Дж., И Чаудхари, Н. (2015). Окситоцин снижает чувствительность к сладкому вкусу у мышей. Physiol. Behav. 141, 103–110. DOI: 10.1016 / j.physbeh.2014.12.048
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Стаматакис, А. М., Ван Свитен, М., Басири, М. Л., Блэр, Г. А., Кантак, П., и Стубер, Г. Д. (2016). Глутаматергические нейроны латеральной области гипоталамуса и их проекции на боковые габенулы регулируют питание и вознаграждение. J. Neurosci. 36, 302–311. DOI: 10.1523 / JNEUROSCI.1202-15.2016
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Стернсон, С. М., Николас Бетли, Дж. И Цао, З. Ф. Х. (2013). Нейронные цепи и мотивационные процессы при голоде. Curr. Opin. Neurobiol. 23, 353–360. DOI: 10.1016 / j.conb.2013.04.006
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Ван, Л., Гиллис-Смит, С., Пэн, Ю., Чжан, Дж., Чен, X., Зальцман, К. Д., и другие. (2018). Кодирование валентности и идентичности во вкусовой системе млекопитающих. Природа 558, 127–131. DOI: 10.1038 / s41586-018-0165-4
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Ву, К., Бойл, М. П., и Палмитер, Р. Д. (2009). Потеря ГАМКергической передачи сигналов нейронами AgRP к парабрахиальному ядру приводит к голоданию. Cell 137, 1225–1234. DOI: 10.1016 / j.cell.2009.04.022
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Ямамото, Т., Оомура, Ю., Нишино, Х., Ау, С., Накано, Ю., и Немото, С. (1984). Активность орбитофронтального нейрона обезьяны во время эмоционального и пищевого поведения. Brain Res. Бык. 12, 441–443. DOI: 10.1016 / 0361-9230 (84)
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Ясумацу, К., Ивата, С., Иноуэ, М., и Ниномия, Ю. (2019). Информация о качестве вкуса жирных кислот через GPR120 на переднем языке мышей. Acta Physiol. 226: e13215. DOI: 10.1111 / apha.13215
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Йошида Р., Ногучи К., Шигемура Н., Джётаки М., Такахаши И., Маргольски Р. Ф. и др. (2015). Лептин подавляет реакцию вкусовых клеток мыши на сладкие соединения. Диабет 64, 3751–3762. DOI: 10.2337 / db14-1462
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Zhang, J., Jin, H., Zhang, W., Ding, C., O’Keeffe, S., Ye, M., et al. (2019). Ощущение кислого ощущения от языка к мозгу. Ячейка 179, 392–402.e15. DOI: 10.1016 / j.cell.2019.08.031
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Чжао, Ф. Л., Шен, Т., Кая, Н., Лу, С. Г., Цао, Ю., и Хернесс, С. (2005). Экспрессия, физиологическое действие и паттерны коэкспрессии нейропептида Y в клетках вкусовых рецепторов крыс. Proc. Natl. Акад. Sci. США 102, 11100–11105. DOI: 10.1073 / pnas.0501988102
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Чжао, Г.Q., Zhang, Y., Hoon, M.A., Chandrashekar, J., Erlenbach, I., Ryba, N.J.P. и др. (2003). Рецепторы сладкого вкуса и вкуса умами млекопитающих. Ячейка 115, 255–266. DOI: 10.1016 / S0092-8674 (03) 00844-4
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Нейронная обработка вкуса
BMC Neurosci. 2007; 8 (Дополнение 3): S5.
1 и 2Christian H Lemon
1 Кафедра анатомии и нейробиологии Научного центра здравоохранения Университета Теннесси, 855 Monroe Ave., Suite 515, Memphis, TN 38163, USA
Donald B Katz
2 Департамент психологии и Национальный центр сложных систем Волена, Университет Брандейса, Волен 208 / MS 013, 415 South St., Waltham, MA 02454, США
1 Департамент анатомии и нейробиологии, Центр медицинских наук Университета Теннесси, 855 Монро Авеню, Люкс 515, Мемфис, TN 38163, США
2 Департамент психологии и Национальный центр сложных систем Волена, Брандейс Университет, Волен 208 / MS 013, 415 South St., Waltham, MA 02454, USA
Автор, ответственный за переписку.Приложение
Химические чувства: последние достижения и новые обещания
Стивен Д. Мангер
Авторские права © 2007 Lemon and Katz; лицензиат BioMed Central Ltd.Это статья в открытом доступе, распространяемая в соответствии с условиями лицензии Creative Commons Attribution License (http://creativecommons.org/licenses/by/2.0), которая разрешает неограниченное использование, распространение и воспроизведение на любом носителе. при условии правильного цитирования оригинала.
Эта статья цитируется в других статьях в PMC.Abstract
Несмотря на то, что в последнее время в области нейробиологии вкуса было много достижений, наши знания о том, как устроена нервная система для обработки информации о вкусе, все еще далеки от полных. Многие исследования по этой теме были сосредоточены на понимании того, как вкусовые нейронные цепи пространственно организованы для представления информации о качестве вкуса (например, «сладкий», «соленый», «горький» и т. Д.). Аргументы, относящиеся к этому вопросу, в основном сосредоточены на том, передаются ли вкусы по выделенным нейронным каналам или по образцу активности в нейронной популяции.Но в настоящее время появляется все больше свидетельств того, что время нейронных событий также может внести важный вклад в представление вкуса. В этом обзоре мы пытаемся обобщить недавние открытия в области, относящейся к этим вопросам. И пространство, и время являются переменными, вероятно, связанными с механизмом вкусового нейронного кода: информация о вкусе, по-видимому, находится в пространственных и временных моделях активации вкусовых нейронов. Более того, организация вкусовой сети в мозгу предполагает, что параметры пространства и времени распространяются на нейронную обработку вкусовой информации в гораздо более широком масштабе.
Введение
В общем, есть две модели пространственного кодирования, которые были предложены для учета нейронного представления информации о вкусе. Одна точка зрения, известная как теория «маркированной линии», предполагает, что нейроны кодируют вкус двоичным образом: когда клетки активны (то есть «включены»), они сигнализируют о наличии определенного признака стимула, в данном случае единственного вкуса. качество [1,2]. Когда эти же нейроны неактивны или «выключены», стимул, который вызывает это конкретное качество, отсутствует.Таким образом, активация клетки служит одной и только одной цели под эгидой теории меченых линий. В противоположность этой точке зрения, некоторые утверждали, что вкус передается паттерном активности популяции нейронов [3,4]. В теории «межнейронного паттерна» отдельные нейроны вносят свой вклад в представление множества качеств стимула, а информация о качестве сигнализируется ответом нейрональной популяции.
Хотя споры о кодировании в значительной степени колебались между тем, использует ли вкус линии или шаблоны, традиционные модели пространственного кодирования упускают из виду информационные зависимости, которые могут присутствовать во времени потенциалов действия или во временных взаимодействиях между вкусовыми нейронами.Однако сама природа организации вкусовых цепей в центральной нервной системе (ЦНС) в виде интерактивных сетей, расположенных последовательно, параллельно и периодически, налагает временную структуру на активность нейронов в любом заданном ядре или области вкуса. Такая структура может выполнять различные функции при обработке вкуса, например, для развития пространственных представлений о вкусовых стимулах во времени, связанных с различными внешними и организменными переменными. Здесь мы суммируем последние разработки, которые проливают новый свет на то, как параметры пространства и времени могут способствовать нейронной обработке информации о вкусе.
Пространственная обработка: вкусовые рецепторы и мозг
В некоторых отношениях механизм маркированных линий, вероятно, наименее сложная форма пространственного кодирования, которую могла бы принять сенсорная нейронная цепь. Интерес к линейному коду как к правдоподобному объяснению работы цепей для определения вкуса был усилен результатами недавних молекулярных и геномных исследований вкусовых рецепторов. Эти исследования выявили два семейства рецепторов, связанных с G-белком, известных как рецепторы T1r и T2r, участвующих в передаче различных вкусовых стимулов.Члены класса T1r объединяются в гетеродимерные функциональные рецепторы, которые воспринимают приятные вкусовые стимулы. В частности, рецептор T1r3 / T1r2 распознает некоторые лиганды, описанные людьми как сладкие на вкус, тогда как рецептор T1r3 / T1r1 участвует в обнаружении аминокислотных стимулов [5,6]. С другой стороны, рецепторы семейства T2r участвуют в обнаружении неприятных, горьких на вкус лигандов [7,8]. Было обнаружено, что эти рецепторы сладких, умами и горьких стимулов экспрессируются в неперекрывающихся подмножествах клеток вкусовых луковиц (ТВП) в эпителии ротовой полости, что было интерпретировано как свидетельство клеточной специфичности по отношению к одному качеству стимула [9-11] .Мыши, сконструированные для экспрессии рецепторов безвкусного соединения в TBC, которые обычно содержат рецепторы сладкого T1r или T2r горького, демонстрируют соответствующее предпочтение или отвращение к этому лиганду [12,13]. Более того, экспрессия горьких рецепторов в «сладких» ТВП T1r приводит к поведенческому влечению к горьким лигандам [12]. Некоторые утверждали, что эти данные указывают на то, что отдельные ОКЧ реагируют на стимулы только одного класса вкусовых качеств и что информация о данном качестве передается по одному из нескольких выделенных, помеченных нейронных каналов [9,12-14].
Хотя неперекрывающиеся паттерны экспрессии рецепторов T1r и T2r рекламировались как свидетельство кодирования маркированной линии, другие данные рисуют иную картину обработки вкуса на периферии. Функциональные исследования с использованием электрофизиологии патч-кламп и методов визуализации кальция показали, что многие ТВЧ в целом чувствительны к стимулам разного вкусового качества, при этом некоторые ТВП реагируют как на сладкие, так и на горькие стимулы [15-17]. Более того, имеются данные о множественных рецепторах для стимулов сладкого и умами [18,19], что делает выводы о периферической обработке этих вкусовых веществ, сделанные на основе исследований отдельных видов рецепторов.Психофизические исследования не обнаружили различий в порогах обнаружения сахарозы или глутамата натрия между мышами, генетически модифицированными с отсутствием рецептора T1r3, и контрольными животными дикого типа [20], предполагая, что T1r3-независимые рецепторы играют важную роль в обнаружении сладких стимулов и стимулов умами. Наконец, теперь есть данные, свидетельствующие о том, что вкусовые клетки обмениваются информацией с соседними клетками внутри зачатка и что существуют отдельные популяции клеток для восприятия вкусовых стимулов и связи с афферентными нервами [21-23].Это повышает вероятность того, что информация от вкусовых рецепторных клеток с различными настраивающими свойствами могла бы сходиться на общих клетках вкусовых рецепторов для передачи в мозг [23]. Обработка во вкусовых сосочках потенциально может запутать интерпретацию данных экспрессии рецепторных генов, показывающих выделенные «линии» для вкусовых качеств. Дальнейшие исследования путей коммуникации внутри вкусовых рецепторов прольют свет на сложности взаимодействия между ТВС.
Вкусовые рецепторы и TBCs участвуют только на самых ранних стадиях обработки вкуса и составляют небольшую часть нейронной массы, участвующей в передаче сигналов и представлении информации о вкусе, которая, очевидно и критически, включает в себя деятельность нейронов и нейронных сетей нижестоящих в Центральная нервная система.Чтобы разгадать логику обработки информации о вкусе, потребуется понимание того, как входные данные от рецепторов обрабатываются цепями ЦНС, что невозможно расшифровать из исследований самих вкусовых рецепторов. Можно интуитивно догадаться, что центральные сети вкусов могут быть настроены любым из множества способов для «кодирования» входных сигналов от рецепторов и создания соответствующих перцептивных или поведенческих выходных данных. Очевидно, мозг должен «знать» стратегию кодирования, используемую этими схемами, чтобы «расшифровать» ввод и произвести соответствующий ответ.Но организация центральных сетей вкусов не может быть эффективно объяснена простым установлением их входных (т. Е. Вкусовых рецепторов) и выходных (т. Е. Поведенческих) отношений. Таким образом, хотя стимуляция ТВС, которые экспрессируют рецепторы сладкого, например, несомненно, приведет к передаче «сладкого сообщения» в мозг, восприятие сладости будет следовать независимо от того, закодирован ли этот сигнал по маркированной линии. с помощью популяционного кода или еще не определенного механизма в мозге (рисунок).
Существует несколько конфигураций центральных вкусовых контуров, которые могут объяснять последствия для восприятия, возникающие после стимуляции определенных вкусовых рецепторов. В гипотетической модели A входные данные от вкусовых рецепторных клеток, которые экспрессируют сладкие рецепторы, кодируются по маркированной линии в центральной нервной системе (ЦНС): информация о сладком стимуле принимается исключительно центральными нейронами, которые реагируют только на сладкое. Тогда центральный «декодер» маркированной линии может знать, что «сладкий» стимул присутствует, когда сладкая «линия» активна.В модели B входные данные от вкусовых рецепторных клеток, которые обнаруживают сладости, распределяются по нейронам и представлены кодом паттерна в ЦНС. Здесь сладкий стимул вызывает уникальный паттерн активации клеток. Центральный декодер паттернов может распознать наличие приятного стимула, зная об этом паттерне. При любой стратегии кодирования стимуляция сладких рецепторных клеток приводит к правильному распознаванию сладкого стимула.
Как организованы центральные нейронные цепи вкуса для кодирования информации о качестве стимула? Поиск ответа на этот вопрос был затруднен повсеместной мультичувствительной природой центральных вкусовых нейронов.То есть многочисленные исследования показали, что центральные сети вкусов состоят из категорий нейронов, которые обычно в целом реагируют на стимулы с различными вкусовыми качествами. В противоречие с гипотезой линии, полученной из исследований паттернов экспрессии рецепторов T1r и T2r, несколько исследований показали, что категории центральных вкусовых нейронов, которые сильно реагируют на сладкие или горькие стимулы, специально не настроены только на эти виды вкусовых добавок. демонстрируя устойчивую чувствительность к раздражителям, таким как соли натрия (т.е., «соленый») и кислый («кислый») растворы [24-27]. Нейрофизиологическое исследование того, как нейроны в ядре единственного тракта (NST) обрабатывают информацию о горьком вкусе, передаваемую нервом VII -го , который обеспечивает входные данные, имеющие решающее значение для поведенческих вкусовых различений [28], показало, что категория нейронов, которые реагировали больше сильно на горькие вещества, такие как хинин, денатония бензоат и папаверин, так же хорошо реагировали на умеренные концентрации натриевых солей NaCl и NaNO 3 [26].С точки зрения кода с маркированными линиями, NaCl управляет этими горькими, наиболее чувствительными нейронами так же эффективно, как многие сильно горькие стимулы, такие как хинин, предсказывают, что NaCl должен вызывать заметное «горькое» ощущение, характерное для хинина. . Однако крысы не делают обобщений в поведении вкусов NaCl и хинина в парадигмах кондиционирования [29,30], что позволяет предположить, что эти стимулы воспринимаются грызунами как независимые. Кроме того, крысы предпочитают умеренные концентрации NaCl, тогда как они явно избегают сверхпороговых концентраций хинина [31].Тем не менее, если мы будем обращать внимание исключительно на продукцию горьких наиболее чувствительных нейронов, что, согласно гипотезе линии, должно позволить нам обнаруживать горький вкус, мы не сможем определить, присутствовал ли NaCl или хинин на вкусовом эпителии. Более недавнее исследование NST показало присутствие клеток, избирательных в отношении веществ, имеющих горький вкус [32]. Но было показано, что большинство чувствительных к горечи нейронов, описанных в [32], получают вкусовые сигналы от IX th нерва, который, как полагают, вносит больший вклад в оромоторные рефлексы, чем на идентификацию вкусовых качеств [28,33].Очевидно, становится трудно согласовать нейронные представления о качественной идентичности веществ, имеющих горький вкус, рассматривая только те нейроны, которые наиболее эффективно реагируют на такие стимулы.
То, что категории или «типы» нейронов являются мультичувствительными, предполагает, что выходные данные любого отдельного класса нейронов могут предоставить только двусмысленную информацию о качестве вкуса [34,35], что имеет значение для того, как центральные вкусовые контуры могут быть организованы для представления информации о дегустаторы.Но прежде чем идти дальше, важно внимательно рассмотреть, что анализ нейронов типов может на самом деле рассказать нам об обработке нейронной информации. В нейрофизиологических исследованиях вкуса было обычным делом использовать в качестве единицы анализа тип нейрона, который отражает совокупный ответ нейронов общей категории. Эти категории обычно определяются путем группирования клеток на основе их наилучшего стимула или с помощью многомерных процедур, которые группируют нейроны на основе сходства их профилей реакции на набор стимулов.Анализы, в которых тип нейрона является основным фактором, по-видимому, предполагают, что это объединенный ответ группы нейронов, с которым мозг должен рассчитывать, чтобы расшифровать входной стимул. Но как мозг объединит активность нейронов общего типа? Нужно ли мозгу заниматься всеми ячейками группы или только частью? Более того, соответствует ли схема объединения, используемая мозгом, классификации нейронов, навязанной экспериментатором? Или мозг просто «считывает» активность вкусовых нейронов на основе отдельных ? Конечно, нет однозначных ответов ни на один из этих вопросов.Например, то, как мозг будет объединять активность вкусовых нейронов в NST, вероятно, будет зависеть от специфики синаптических связей между этими клетками и подчиненными нейронами в парабрахиальном ядре, тема, которая не совсем понятна. Кроме того, средний ответ нейронного типа потенциально может недооценивать или переоценивать настраивающие свойства отдельных клеток. Таким образом, оценка эффективности кодирования вкусовых нейронов, вероятно, лучше всего проиндексировать через понимание пределов обработки информации отдельных клеток , которые также будут влиять на эффективность обнаружения стимулов, которая может быть достигнута путем объединения их действий каким-либо образом.Определение этих пределов требует знания того, насколько надежно отдельные вкусовые нейроны реагируют на стимулы с течением времени и в разных исследованиях. Эта тема получила лишь скудное внимание в литературе (но см. [36]).
Так же важно, как изменчивость ответа внутри нейрона, необходимо учитывать продолжительность времени, в течение которого измеряются вкусовые реакции. Многие исследования обработки вкуса позволили количественно оценить активность вкусовых нейронов на основе количества спайков, измеренных в течение 5 или 10 секундных окон «стимул-ответ».Тем не менее, важно признать, что этот период чрезвычайно долгий по сравнению с количеством времени, которое требуется нервной системе, чтобы прийти к перцепционному суждению о качестве вкусовых стимулов. Поведенческие исследования с использованием процедур условного избегания показали, что крысы могут распознавать реакцию и на вкусовые стимулы менее чем через 1 секунду после контакта [37-39]. Это означает, что необходимая и достаточная информация о качестве стимула заложена в пиковой активности вкусовых нейронов в течение первых нескольких сотен миллисекунд вызванной активности.Это краткое окно, содержащее важную информацию об идентичности стимула, может соответствовать только нескольким или нескольким максимумам потенциалов действия во многих вкусовых нейронах, когда они находятся под воздействием вкусового влечения.
Принимая во внимание эти вопросы, в недавнем нейрофизиологическом исследовании использовалась теоретическая методика, примененная к отдельным нейронам NST крысы, чтобы изучить, как изменчивость частоты всплесков отдельных клеток в течение первой секунды обработки стимула может влиять на способность скорости всплеска предсказывать стимул. идентифицировать [40].Во всех протестированных нейронах было обнаружено, что стимулы с разными вкусовыми качествами производили переменные и перекрывающиеся распределения скорости спайков до такой степени, что реакция отдельного нейрона сразу после контакта со стимулом была ненадежным индикатором качества стимула. Этот вывод предполагает, что может оказаться трудным, если не невозможным, декодировать ввод стимула во время этого окна, прикрепляя сообщения вкуса к выделенным нейронам или их группам и просто сообщая сообщение, назначенное блоку обработки при активации.Дальнейший анализ, проведенный в [40], показал, что разные вкусовые вещества вызывают уникальные относительные пиковые отношения между несколькими нейронами NST, сравниваемыми параллельно. «Читатель», который обращает внимание на эту информацию и знает стимул, связанный с каждым паттерном взаимоотношений, может, в принципе, вычислить различия между различными вкусовыми агентами (рисунок). Эти данные представляют собой разумную модель того, как вкусовая система может использовать пространственное кодирование для вычисления качества стимула: вместо того, чтобы приписывать значение отдельным нейронам или их категориям, центральные вкусовые контуры могут передавать информацию о качестве через относительную активность нескольких нейронов параллельно.Другие данные также подтверждают это мнение [4,34,35].
Стимулы с разными вкусовыми качествами создают уникальные паттерны относительного возбуждения центральных вкусовых нейронов в течение первой секунды обработки стимула. Здесь сравнивались уровни пиков пероральной стимуляции сахарозой (прототип «сладкого» стимула), NaCl («соленый»), HCl («кислый») или хинин («горький») между вкусовыми нейронами, записанными от крысиного NST с использованием теоретический метод, основанный на теории статистических решений.Эта модель зависит от того, работают ли разные клетки с одинаковой или достоверно разной скоростью спайков, когда они находятся под действием определенного стимула. Результат этого анализа применительно ко всем возможным парам нейронов среди шести случайно выбранных ячеек представлен графически в виде набора полуматриц. Затемненный матричный элемент представляет, что нейрон i th (обозначенный вдоль столбцов матрицы) конкретной пары активизировался с заметно большей скоростью, чем j th (строки).Незаштрихованный элемент означает одинаковую частоту всплесков (не различную) между нейронами, тогда как полутоновое затенение указывает, что j th сработал заметно быстрее, чем i th . Можно видеть, что разные стимулы вызывают уникальные отношения относительного ответа между этими клетками. Нижестоящий процессор этих нейронов со знанием стимула, связанного с каждым ответным отношением, может, в принципе, вычислить различия между этими стимулами.Перепечатано из [40] с разрешения журнала Journal of Neuroscience .
Хотя интригующая модель, представленная в [40] (рисунок), представляет собой описание того, как пространственное нейронное представление может потенциально работать со вкусом. Модель показывает, что информация о вкусовой идентичности может быть извлечена гипотетическим читателем, который сравнивает нейроны NST в рамках теоретической основы. Но, конечно, неизвестно, примет ли нервная система аналогичный алгоритм для регистрации вкусовой идентичности.Понимание того, как именно вкусовые нейроны «считываются» мозгом, потребует знания архитектуры сетей, связывающих эти клетки с нижележащими нейронами и ядрами, а также функций передачи информации, используемых в этих цепях. Детали здесь еще предстоит проработать.
Время и интерактивная обработка по вкусу
Исторически большинство моделей вкусового кодирования не обращали внимания на информацию, которая могла бы передаваться зависимостями от времени нейронных событий.Фактически, строгий межнейронный паттерн и модели с маркированными линиями предполагают, что время имеет очень мало значения для «вкусового кодирования». Однако свидетельств, подтверждающих это предположение, на самом деле немного. Использование средних значений скорости возбуждения 5–10 секунд по большей части было принято по необходимости, а не по замыслу, поскольку вкусовые нейроны часто удерживаются только посредством однократного предъявления индивидуальных стимулов. Хотя в некоторых исследованиях сообщается о корреляциях между общей частотой возбуждения и вкусовым поведением членов большой батареи стимулов, эти корреляции в лучшем случае умеренные и описывают лишь широкое сходство между вкусами [41].Кроме того, часто цитируемый факт, что грызуны могут при некоторых обстоятельствах демонстрировать распознавание вкуса за ~ 200 мс [39,42], не может служить серьезным обвинением против временного кодирования по крайней мере по двум причинам: 1) этот результат вызывает одинаковые проблемы для всех текущих схем кодирования — учитывая, что информация о вкусе поступает в NST относительно медленно [43] и что вкусовые реакции являются относительно низкоскоростным феноменом, нейроны имеют возможность вызвать лишь несколько всплесков в первые 200 мсек. вкус на языке животного — недостаток информации для надежного распознавания активированного нейрона или пространственного паттерна; и 2) многие виды поведения, связанные со вкусом, происходят только в масштабе времени, на порядок превышающем тот, который описан в вышеупомянутых исследованиях [44] — создаваемый код зависит от состояния внимания животного [45], наряду с многие другие переменные для конкретных задач.
Между тем, есть несколько причин учитывать время при кодировании вкуса, помимо того факта, что различное вкусовое поведение требует разного количества времени обработки стимула. Во-первых, сети в NST [46,47] и за его пределами, включая более крупные сети соединений с прямой связью и обратной связью [48-51], почти гарантируют, что обработка вкуса и кодирование будут модулироваться во времени, поскольку нейроны получают асинхронный ввод из нескольких источников. (к этой теме мы вернемся в ближайшее время).Кроме того, вполне вероятно, что кодирование вкуса имеет временной аспект, потому что большинство других сенсорных реакций, как было показано, имеют такой же аспект [52-54].
То немногое, что было собрано до сих пор, предполагает, что вкусовые нейроны действительно реагируют на вкусы с изменяющимся во времени паттерном активности как на уровне ствола мозга [36], так и на уровне коры головного мозга [55,56]. В некоторой степени этот результат получается потому, что сбор данных нескольких испытаний, который требуется для комплексного анализа, выявляет тонкие, поэтапные и многоэтапные ответы, которые упускаются при анализе общей скорости отдельных наборов данных испытаний.Такие наборы данных также показывают, что на профили ответов, определенные в экспериментах, в которых каждый вкус был доставлен только один или два раза, часто чрезмерно влияет изменчивость реакции от испытания к испытанию [36], которой печально известны нейроны ЦНС. Таким образом, зависимость от сильных тонических реакций заставляет исследователей неверно трактовать кодирование вкуса как в том, что наблюдается, так и в том, что упускается.
Сторонники гипотез пространственного кодирования считают более тонкие, изменяющиеся во времени модуляции, наблюдаемые во вкусовых реакциях, либо «шумом», либо «несущественными».«Таким выводам, однако, противоречат по крайней мере два типа исследований: те, которые демонстрируют, что временные коды несут конкретную полезную информацию, и те, которые показывают, что животные могут делать вкусовые суждения, основанные исключительно на временных кодах.
Что касается первого из них. В типах исследований вкусовые коды, записанные у бодрствующих животных, не просто меняются во времени — они, по-видимому, «мультиплексируют» информацию [55,57], что также было показано как для зрительной [58], так и для обонятельной реакции [59]. , ранние части вкусовых реакций, по крайней мере, в коре головного мозга, несут информацию о качестве вкуса, тогда как более поздние части несут информацию о вкусовых качествах.Более свежие данные показывают, что изменения в ответах «поздней фазы» конкретно связаны с изменениями вкусовых качеств, измеряемых с точки зрения орофациального поведения [45]. Это служит доказательством того, что временное кодирование вкуса может отражать обработку вкуса, когда животное решает, что оно думает о вкусе.
Более того, изучение ансамблевых ответов показывает, что то, что в записях отдельных нейронов является случайной изменчивостью от испытания к испытанию, на самом деле согласовано на уровне популяции.Когда всплески и изменения частоты импульсов в корковых нейронах связаны с всплесками и изменениями в других, одновременно регистрируемых нейронах (а не с началом стимула), можно увидеть, что они прогрессируют через серию состояний, зависящих от вкуса, которые «развиваются» с разной скоростью в разных испытаниях [60]. Последовательность состояний предоставляет значительно лучшую информацию о вкусе, полученном в конкретном испытании, чем методы, основанные на общих показателях (или даже изменяющихся во времени PSTH). Это и предыдущие исследования показывают, что временные коды важны для понимания обработки вкусов.Хотя ясно, что зазоры и облизывание могут быть произведены цепями центрального генератора паттернов ствола мозга (CPG), исследования повреждений показывают, что большая сеть областей переднего мозга — по крайней мере, префронтальная кора, миндалина и гипоталамус — отвечает за принятие решений о вкусовых качествах. [61-63], и, таким образом, для решения, какие паттерны ствола мозга образуются у интактного животного; в качестве аналогии подумайте о контроле ходьбы, который может осуществляться спинализированными млекопитающими, и все же по своей сути включает мозжечковые и корковые механизмы контроля [64].
Но, конечно, для абсолютного доказательства требуются эксперименты, показывающие, что используются временные коды, а не просто то, что их можно использовать. В этом отношении интересны два недавних исследования. В одном из них исследователи электрически стимулировали NTS крыс, используя временную структуру спайков, которая была ранее записана в ответ на вкус, когда эти крысы пили воду. Крысы реагировали на воду, как если бы она была горькой при стимуляции хинином [65]. Более того, условное отвращение к сахарозе распространено на воду, потребляемую одновременно с доставкой сахарозы, но не на доставку других вкусовых моделей.Эти данные убедительно свидетельствуют о том, что крысы используют временные коды для вкуса.
Гусеницы Manduca , по-видимому, также используют временную информацию при определении вкусов [66]. У этих существ есть несколько трансдуктивных элементов (сенсилл), которые реагируют на широкий спектр горьких раздражителей; эти сенсиллы реагируют на аристолохиновую кислоту (АК) учащающимися всплесками, а на кофеин — замедляющимися всплесками. Когда прилагаются усилия к тому, чтобы уравнять общие пиковые реакции на эти стимулы (т.е., пространственные коды), гусеницы по-прежнему различают АК и салицин (еще один горький вкус, который вызывает замедление нервной реакции), но не между кофеином и салицином. Эти данные убедительно подтверждают ценность времени во вкусе.
Активность нейронных сетей обеспечивает вероятный механизм для большинства явлений временного кодирования, описанных выше. Исследования, в которых использовались многоэлектродные записи для выявления взаимодействий между вкусовыми нейронами [67-70], и те, в которых использовалась комбинация стимуляции / инактивации и записи для выявления влияния обратной связи на вкусовые реакции NTS и PbN [48-51], демонстрируют реальность сетевой обработки во вкусе.Ясно, что вкусовые нейроны «разговаривают» друг с другом, и этот разговор идет между различными нейронами в пределах отдельных областей мозга, в конвергенции нейронов с несопоставимыми паттернами ответа на единичные нижестоящие мишени и в модуляции основных ответов нейронами переднего мозга, несущими более высокий уровень. обработанная информация. Почти неизбежно, что нейронные взаимодействия вызовут модуляцию вкусовых реакций во времени.
Есть свидетельства того, что «код» вкуса в мозге может включать как пространственные [71,72], так и временные аспекты активности вкусовых нейронов.Но параметры пространства и времени также имеют решающее значение для вкусового кодирования в гораздо большем масштабе. Обработка вкуса — это событие сетевого уровня, в котором участвуют распределенные структуры CNS, которые взаимодействуют друг с другом в зависимости от времени. Именно эта интерактивная обработка между узлами вкусовой системы регулирует информационный поток через центральный нейраксис вкуса, в конечном итоге формируя и развивая нейронный «код» вкуса относительно различных параметров текущей перцептивной и поведенческой обработки.Понимание таких нейронных взаимодействий могло бы открыть убедительное окно для организации цепей на вкус, хотя наши знания здесь все еще находятся в зачаточном состоянии.
Конкурирующие интересы
Авторы заявляют, что у них нет конкурирующих интересов.
Вклад авторов
C.H.L. и Д.Б.К. разработал план этой рукописи. C.H.L. написал реферат, введение, раздел «Пространственная обработка: вкусовые рецепторы и мозг» и заключительный абзац.D.B.K. написал раздел «Время и интерактивная обработка во вкусе». C.H.L и D.B.K. отредактировал полную рукопись.
Ссылки
- Pfaffmann C, Frank M, Bartoshuk LM, Snell TC. Кодирование вкусовой информации в chorda tympani беличьей обезьяны. В: Sprague JM, Epstein AN, редактор. Прогресс в психобиологии и физиологической психологии. Vol. 6. Нью-Йорк: Academic Press; 1976. С. 1-27. [Google Scholar]
- Хеллекант Г., Ниномия Ю., Данилова В. Вкус шимпанзе.III: маркированная линия в сладком вкусе. Physiol Behav. 1998. 65: 191–200. DOI: 10.1016 / S0031-9384 (97) 00532-5. [PubMed] [CrossRef] [Google Scholar]
- Пфаффманн К. Афферентный код сенсорного качества. Am Psychol. 1959; 14: 226–232. DOI: 10,1037 / h0049324. [CrossRef] [Google Scholar]
- Эриксон Р.П. Сенсорные нейронные паттерны и вкусовые ощущения. В: Зоттерман Ю., редактор. Обоняние и вкус. Vol. 1. Оксфорд: Pergamon Press; 1963. С. 205–213. [Google Scholar]
- Nelson G, Chandrashekar J, Hoon MA, Feng L, Zhao G, Ryba NJ, Zuker CS.Аминокислотный рецептор вкуса. Природа. 2002; 416: 199–202. DOI: 10,1038 / природа726. [PubMed] [CrossRef] [Google Scholar]
- Li X, Staszewski L, Xu H, Durick K, Zoller M, Adler E. Человеческие рецепторы сладкого вкуса и вкуса умами. Proc Natl Acad Sci USA. 2002; 99: 4692–4696. DOI: 10.1073 / pnas.0720. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
- Чандрашекар Дж., Мюллер К.Л., Хун М.А., Адлер Э., Фенг Л., Го В., Цукер С.С., Рыба, штат Нью-Джерси. T2R действуют как рецепторы горького вкуса. Клетка. 2000; 100: 703–711.DOI: 10.1016 / S0092-8674 (00) 80706-0. [PubMed] [CrossRef] [Google Scholar]
- Bufe B, Hofmann T, Krautwurst D, Raguse JD, Meyerhof W. Рецептор TAS2R16 человека опосредует горький вкус в ответ на бета-глюкопиранозиды. Нат Жене. 2002; 32: 397–401. DOI: 10,1038 / нг1014. [PubMed] [CrossRef] [Google Scholar]
- Скотт К. Сладкий и горький вкус млекопитающих. Curr Opin Neurobiol. 2004. 14: 423–427. DOI: 10.1016 / j.conb.2004.06.003. [PubMed] [CrossRef] [Google Scholar]
- Nelson G, Hoon MA, Chandrashekar J, Zhang Y, Ryba NJ, Zuker CS.Рецепторы сладкого вкуса млекопитающих. Клетка. 2001; 106: 381–390. DOI: 10.1016 / S0092-8674 (01) 00451-2. [PubMed] [CrossRef] [Google Scholar]
- Adler E, Hoon MA, Mueller KL, Chandrashekar J, Ryba NJP, Zuker CS. Новое семейство вкусовых рецепторов млекопитающих. Клетка. 2000; 100: 693–702. DOI: 10.1016 / S0092-8674 (00) 80705-9. [PubMed] [CrossRef] [Google Scholar]
- Мюллер К.Л., Хун М.А., Эрленбах И., Чандрашекар Дж., Цукер С.С., Рыба, штат Нью-Джерси. Рецепторы и логика кодирования горечи. Природа. 2005. 434: 225–229.DOI: 10,1038 / природа03352. [PubMed] [CrossRef] [Google Scholar]
- Zhao GQ, Zhang Y, Hoon MA, Chandrashekar J, Erlenbach I., Ryba NJ, Zuker CS. Рецепторы сладкого вкуса и вкуса умами млекопитающих. Клетка. 2003. 115: 255–266. DOI: 10.1016 / S0092-8674 (03) 00844-4. [PubMed] [CrossRef] [Google Scholar]
- Zhang Y, Hoon MA, Chandrashekar J, Mueller KL, Cook B., Wu D, Zuker CS, Ryba NJ. Кодирование сладкого, горького и умами вкусов: разные рецепторные клетки имеют сходные сигнальные пути. Клетка.2003; 112: 293–301. DOI: 10.1016 / S0092-8674 (03) 00071-0. [PubMed] [CrossRef] [Google Scholar]
- Сато Т., Бейдлер Л.М. Широкая настройка вкусовых клеток крысы на четыре основных вкусовых стимула. Chem Senses. 1997. 22: 287–293. DOI: 10.1093 / chemse / 22.3.287. [PubMed] [CrossRef] [Google Scholar]
- Gilbertson TA, Boughter JD, Jr, Zhang H, Smith DV. Распределение вкусовой чувствительности во вкусовых клетках крыс: ответы целых клеток на апикальную химическую стимуляцию. J Neurosci. 2001; 21: 4931–4941. [Бесплатная статья PMC] [PubMed] [Google Scholar]
- Кайседо А., Ким К.Н., Ропер С.Д.Отдельные вкусовые клетки мыши реагируют на несколько химических раздражителей. J Physiol. 2002; 544: 501–509. DOI: 10.1113 / jphysiol.2002.027862. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
- Маруяма Ю., Перейра Э., Маргольски РФ, Чаудхари Н., Ропер С.Д. Ответы умами во вкусовых клетках мыши указывают на более чем один рецептор. J Neurosci. 2006; 26: 2227–2234. DOI: 10.1523 / JNEUROSCI.4329-05.2006. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
- Damak S, Rong M, Yasumatsu K, Kokrashvili Z, Varadarajan V, Zou S, Jiang P, Ninomiya Y, Margolskee RF.Обнаружение сладкого вкуса и вкуса умами при отсутствии вкусового рецептора T1r3. Наука. 2003. 301: 850–853. DOI: 10.1126 / science.1087155. [PubMed] [CrossRef] [Google Scholar]
- Delay ER, Эрнандес Н.П., Бромли К., Маргольски РФ. Вкусовые пороги сахарозы и глутамата натрия и способность к различению мышей с нокаутом T1R3. Chem Senses. 2006. 31: 351–357. DOI: 10,1093 / chemse / bjj039. [PubMed] [CrossRef] [Google Scholar]
- Zhao FL, Shen T, Kaya N, Lu SG, Cao Y, Herness S. Экспрессия, физиологическое действие и коэкспрессия нейропептида Y в клетках вкусовых рецепторов крыс.Proc Natl Acad Sci USA. 2005. 102: 11100–11105. DOI: 10.1073 / pnas.0501988102. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
- Kaya N, Shen T, Lu SG, Zhao FL, Herness S. Паракринная сигнальная роль серотонина во вкусовых рецепторах крыс: экспрессия и локализация подтипов серотониновых рецепторов . Am J Physiol Regul Integr Comp Physiol. 2004; 286: R649–658. [PubMed] [Google Scholar]
- ДеФазио Р.А., Дворянчиков Г., Маруяма Ю., Ким Дж. У., Перейра Е., Ропер С. Д., Чаудхари Н. Отдельные популяции рецепторных клеток и пресинаптических клеток во вкусовых сосочках мыши.J Neurosci. 2006; 26: 3971–3980. DOI: 10.1523 / JNEUROSCI.0515-06.2006. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
- Verhagen JV, Giza BK, Scott TR. Влияние амилорида на вкусовые реакции вентропостеромедиального ядра таламуса у крыс. J Neurophysiol. 2005. 93: 157–166. DOI: 10.1152 / jn.00823.2003. [PubMed] [CrossRef] [Google Scholar]
- Ди Лоренцо П.М., Лимонный чемпионат, Рейх CG. Динамическое кодирование вкусовых стимулов в стволе мозга: влияние коротких импульсов вкусовых стимулов на последующие вкусовые реакции.J Neurosci. 2003; 23: 8893–8902. [Бесплатная статья PMC] [PubMed] [Google Scholar]
- Lemon CH, Smith DV. Нервное представление горького вкуса в ядре единственного тракта. J Neurophysiol. 2005; 94: 3719–3729. DOI: 10.1152 / jn.00700.2005. [PubMed] [CrossRef] [Google Scholar]
- Scott TR, Giza BK. Каналы кодирования во вкусовой системе крысы. Наука. 1990; 249: 1585–1587. DOI: 10.1126 / science.2171145. [PubMed] [CrossRef] [Google Scholar]
- St John SJ, Spector AC. Поведенческое различие между хинином и KCl зависит от сигнала седьмого черепного нерва: это влияет на функциональную роль вкусовых нервов у крыс.J Neurosci. 1998. 18: 4353–4362. [Бесплатная статья PMC] [PubMed] [Google Scholar]
- Morrison GR. Паттерны поведенческой реакции крыс на солевые раздражители. Может J Психология. 1967; 21: 141–152. [Google Scholar]
- Nowlis GH, Frank ME, Pfaffmann C. Специфика приобретенного отвращения к вкусовым качествам у хомяков и крыс. J Comp Physiol Psychol. 1980; 94: 932–942. DOI: 10,1037 / h0077809. [PubMed] [CrossRef] [Google Scholar]
- Пфаффманн К. Вкус, его сенсорные и мотивирующие свойства.Am Sci. 1964; 52: 187–206. [Google Scholar]
- Geran LC, Travers SP. Одиночные нейроны в ядре солитарного тракта избирательно реагируют на раздражители горького вкуса. J Neurophysiol. 2006; 96: 2513–2527. DOI: 10.1152 / jn.00607.2006. [PubMed] [CrossRef] [Google Scholar]
- Трэверс Дж. Б., Гриль Х. Дж., Норгрен Р. Влияние перерезаний языкоглоточного и барабанного нерва на прием и отторжение мозговых стимулов: электромиографический анализ на крысах. Behav Brain Res. 1987. 25: 233–246.DOI: 10.1016 / 0166-4328 (87)
-4. [PubMed] [CrossRef] [Google Scholar] - Smith DV, St John SJ. Нейронное кодирование вкусовой информации. Curr Opin Neurobiol. 1999; 9: 427–435. DOI: 10.1016 / S0959-4388 (99) 80064-6. [PubMed] [CrossRef] [Google Scholar]
- Scott TR, Giza BK. Проблемы вкусового нейронного кодирования: где они стоят сегодня. Physiol Behav. 2000; 69: 65–76. DOI: 10.1016 / S0031-9384 (00) 00189-X. [PubMed] [CrossRef] [Google Scholar]
- Di Lorenzo PM, Victor JD. Вариабельность вкусовых реакций и временное кодирование в ядре единственного тракта крысы.J Neurophysiol. 2003; 90: 1418–1431. DOI: 10.1152 / jn.00177.2003. [PubMed] [CrossRef] [Google Scholar]
- Скотт TR. Поведенческая поддержка теории нервного вкуса. Physiol Behav. 1974; 12: 413–417. DOI: 10.1016 / 0031-9384 (74)
-8. [PubMed] [CrossRef] [Google Scholar]
- Halpern BP. Время как фактор вкуса: временные паттерны вкусовой стимуляции и реакции. В: Pfaff DW, редактор. Вкус, обоняние и центральная нервная система. Нью-Йорк, Нью-Йорк: издательство Рокфеллерского университета; 1985 г.С. 181–209. [Google Scholar]
- Halpern BP, Tapper DN. Вкусовые стимулы: время качественного кодирования. Наука. 1971; 171: 1256–1258. DOI: 10.1126 / science.171.3977.1256. [PubMed] [CrossRef] [Google Scholar]
- Lemon CH, Smith DV. Влияние вариабельности ответа на кодирование центральных вкусовых нейронов. J Neurosci. 2006; 26: 7433–7443. DOI: 10.1523 / JNEUROSCI.0106-06.2006. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
- Накамура К., Норгрен Р. Вкусовые ответы нейронов в ядре единственного тракта бодрствующих крыс: расширенный массив стимулов.J Neurophysiol. 1993; 70: 879–891. [PubMed] [Google Scholar]
- Boughter JD, Jr, John SJ, Noel DT, Ndubuizu O, Smith DV. Краткосрочный тест на горький вкус у мышей. Chem Senses. 2002. 27: 133–142. DOI: 10.1093 / chemse / 27.2.133. [PubMed] [CrossRef] [Google Scholar]
- Эриксон Р.П., Ди Лоренцо П.М., Вудбери, Массачусетс. Классификация вкусовых реакций в стволе мозга: принадлежность к нечетким множествам. J Neurophysiol. 1994; 71: 2139–2150. [PubMed] [Google Scholar]
- Halpern BP. Временные характеристики вкусовых суждений человека как калибровка вкусовых, связанных с событием потенциалов и вкусовых магнитоэнцефалографов.Chem Senses. 2005; 30: i228 – i229. DOI: 10,1093 / chemse / bjh297. [PubMed] [CrossRef] [Google Scholar]
- Fontanini A, Katz DB. Зависимая от состояния модуляция изменяющихся во времени вкусовых реакций. J Neurophysiol. 2006; 96: 3183–3193. DOI: 10.1152 / jn.00804.2006. [PubMed] [CrossRef] [Google Scholar]
- Брэдли Р.М., Грабаускас Г. Нервные цепи на вкус: возбуждение, торможение и синаптическая пластичность в ростральной вкусовой зоне ядра единственного тракта. Ann N Y Acad Sci. 1998. 855: 467–474.DOI: 10.1111 / j.1749-6632.1998.tb10607.x. [PubMed] [CrossRef] [Google Scholar]
- Lemon CH, Di Lorenzo PM. Влияние электрической стимуляции нерва барабанной хорды на вкусовые реакции в ядре единственного тракта. J Neurophysiol. 2002; 88: 2477–2489. DOI: 10.1152 / jn.00094.2002. [PubMed] [CrossRef] [Google Scholar]
- Ли К.С., Чо Ю.К., Смит Д.В. Модуляция парабрахиальных вкусовых нейронов с помощью электрической и химической стимуляции бокового гипоталамуса и миндалины. J Neurophysiol.2005; 93: 1183–1196. DOI: 10.1152 / jn.00828.2004. [PubMed] [CrossRef] [Google Scholar]
- Ли К.С., Чо Ю.К., Смит Д.В. Вкусовые реакции нейронов в одиночном ядре хомяка модулируются центральным ядром миндалины. J Neurophysiol. 2002; 88: 2979–2992. DOI: 10.1152 / jn.00239.2002. [PubMed] [CrossRef] [Google Scholar]
- Lundy RF, Jr, Norgren R. Активность в гипоталамусе, миндалине и коре головного мозга генерирует двустороннюю и конвергентную модуляцию вкусовых нейронов моста. J Neurophysiol.2004. 91: 1143–1157. DOI: 10.1152 / jn.00840.2003. [PubMed] [CrossRef] [Google Scholar]
- Чо Ю.К., Ли С.С., Смит Д.В. Нисходящие влияния латерального гипоталамуса и миндалевидного тела сходятся на нейроны медуллярного вкуса. Chem Senses. 2003. 28: 155–171. DOI: 10.1093 / chemse / 28.2.155. [PubMed] [CrossRef] [Google Scholar]
- Ghazanfar AA, Nicolelis MAL. Пространственно-временной континуум в сенсорных путях млекопитающих. В: Сидни М.Р., редактор. Время и мозг. Австралия: Harwood Press; 2000. С.97–130. [Google Scholar]
- Лоран Г., Стопфер М., Фридрих Р. В., Рабинович М. И., Волковский А., Абарбанель HD. Кодирование запаха как активный, динамичный процесс: эксперименты, вычисления, теория. Annu Rev Neurosci. 2001; 24: 263–297. DOI: 10.1146 / annurev.neuro.24.1.263. [PubMed] [CrossRef] [Google Scholar]
- McClurkin JW, Optican LM, Richmond BJ, Gawne TJ. Параллельная обработка и сложность закодированных во времени нейронных сообщений в визуальном восприятии. Наука. 1991; 253: 675–677. DOI: 10.1126 / наука.18. [PubMed] [CrossRef] [Google Scholar]
- Кац Д.Б., Саймон С.А., Николелис М.А. Динамические и мультимодальные ответы вкусовых нейронов коры у бодрствующих крыс. J Neurosci. 2001; 21: 4478–4489. [Бесплатная статья PMC] [PubMed] [Google Scholar]
- Кобаякава Т., Огава Х., Канеда Х., Аябе-Канамура С., Эндо Х., Сайто С. Пространственно-временной анализ корковой активности, вызванной вкусовой стимуляцией у людей. Химические чувства. 1999; 24: 201–209. DOI: 10.1093 / chemse / 24.2.201. [PubMed] [CrossRef] [Google Scholar]
- Katz DB.Множество разновидностей временного кодирования во вкусовой коре. Chem Senses. 2005; 30: i80 – i81. DOI: 10,1093 / chemse / bjh223. [PubMed] [CrossRef] [Google Scholar]
- Sugase Y, Yamane S, Ueno S, Kawano K. Глобальная и точная информация, кодируемая отдельными нейронами в височной зрительной коре. Природа. 1999; 400: 869–873. DOI: 10,1038 / 23703. [PubMed] [CrossRef] [Google Scholar]
- Friedrich RW, Habermann CJ, Laurent G. Мультиплексирование с использованием синхронизации в обонятельной луковице рыбок данио. Nat Neurosci. 2004; 7: 862–871.DOI: 10,1038 / NN1292. [PubMed] [CrossRef] [Google Scholar]
- Джонс Л.М., Фонтанини А., Кац ДБ. Ансамблевые ответы вкусовых кортикальных нейронов точно предсказывают вкусовые качества. Chem Senses. 2006; 31: A115. [Google Scholar]
- Тузани К., Тагзути К., Велли Л. Повышение отталкивающей ценности вкусовых стимулов после поражения иботеновой кислотой центрального миндалевидного ядра у крысы. Поведенческие исследования мозга. 1997. 88: 133–142. DOI: 10.1016 / S0166-4328 (96) 02273-5. [PubMed] [CrossRef] [Google Scholar]
- Gutierrez R, Carmena JM, Nicolelis MA, Simon SA.Активность орбитофронтального ансамбля отслеживает лизание и выделяет естественные награды. J Neurophysiol. 2006. 95: 119–133. DOI: 10.1152 / jn.00467.2005. [PubMed] [CrossRef] [Google Scholar]
- Ферссиви А., Кардо Б., Велли Л. Пороги неприятия вкусовых предпочтений у крыс повышаются из-за поражения иботеновой кислотой бокового гипоталамуса у крыс. Brain Res. 1987. 437: 142–150. DOI: 10.1016 / 0006-8993 (87)
-6. [PubMed] [CrossRef] [Google Scholar]
- Орловский Г.Н., Делягина Т.Г., Грилнер С.Нейронный контроль передвижений от моллюска к человеку. Оксфорд: издательство Оксфордского университета; 1999. [Google Scholar]
- Ди Лоренцо П.М., Хэллок Р.М., Кеннеди Д.П. Временное кодирование ощущений: имитация вкусовых качеств с помощью электрической стимуляции мозга. Behav Neurosci. 2003. 117: 1423–1433. DOI: 10.1037 / 0735-7044.117.6.1423. [PubMed] [CrossRef] [Google Scholar]
- Глендиннинг Дж. И., Дэвис А., Рай М. Временное кодирование опосредует распознавание насекомыми раздражителей «горького» вкуса. J Neurosci.2006. 26: 8900–8. DOI: 10.1523 / JNEUROSCI.2351-06.2006. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
- Адачи М., Охима Т., Ямада С., Сато Т. Анализ взаимной корреляции пар вкусовых нейронов в ядре единственного тракта крысы. J Neurophysiol. 1989; 62: 501–509. [PubMed] [Google Scholar]
- Кац Д., Саймон С., Николелис М.А.Л. Вкусовые ансамбли нейронов во вкусовой коре головного мозга бодрствующих крыс. J Neurosci. 2002; 22: 1850–1857. [Бесплатная статья PMC] [PubMed] [Google Scholar]
- Yokota T, Satoh T.Трехмерная оценка распределения и размера предполагаемых функциональных единиц во вкусовой коре крыс по межнейронному расстоянию между двумя нейронами с соответствующей активностью. Brain Res Bull. 2001; 54: 575–584. DOI: 10.1016 / S0361-9230 (01) 00464-6. [PubMed] [CrossRef] [Google Scholar]
- Ямада С., Охшима Т., Ода Х., Адачи М., Сато Т. Синхронная разрядка вкусовых нейронов, записанная одновременно в парабрахиальном ядре крысы. J Neurophysiol. 1990; 63: 294–302. [PubMed] [Google Scholar]
- Йошимура Х., Сугай Т., Фукуда М., Сегами Н., Онода Н.Корковые пространственные аспекты оптических внутренних сигналов в ответ на стимулы сахарозы и NaCl. Нейроотчет. 2004; 15: 17–20. DOI: 10.1097 / 00001756-200401190-00005. [PubMed] [CrossRef] [Google Scholar]
- Accolla R, Bathellier B, Petersen CC, Carleton A. Дифференциальное пространственное представление модальностей вкуса во вкусовой коре головного мозга крыс. J Neurosci. 2007. 27: 1396–1404. DOI: 10.1523 / JNEUROSCI.5188-06.2007. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
Синтез и функции ретиноевой кислоты в раннем эмбриональном развитии | Cell & Bioscience
Tang XH, Gudas LJ: Ретиноиды, рецепторы ретиноевой кислоты и рак. Анну Рев Патол. 2011, 6: 345-364. 10.1146 / annurev-pathol-011110-130303
КАС PubMed Статья Google Scholar
Duriancik DM, Lackey DE, Hoag KA: Витамин А как регулятор антигенпрезентирующих клеток. J Nutr. 140 (8): 1395-1399.
Gudas LJ, Wagner JA: Ретиноиды регулируют дифференцировку стволовых клеток. J. Cell Physiol.226 (2): 322-330.
Hogarth CA, Griswold MD: Ключевая роль витамина А в сперматогенезе. J Clin Invest. 120 (4): 956-962.
Мора Дж. Р., Ивата М., фон Андриан У. Х .: Влияние витаминов на иммунную систему: витамины A и D занимают центральное место. Nat Rev Immunol. 2008, 8 (9): 685-698. 10.1038 / nri2378
PubMed Central CAS PubMed Статья Google Scholar
Мукерджи С., Дате А., Патравале В., Кортинг Х.С., Рёдер А., Вайндл Г.: Ретиноиды в лечении старения кожи: обзор клинической эффективности и безопасности.Clin Interv Aging. 2006, 1 (4): 327-348. 10.2147 / ciia.2006.1.4.327
PubMed Central CAS PubMed Статья Google Scholar
Пино-Лагос К., Бенсон М.Дж., Ноэль Р.Дж.: Ретиноевая кислота в иммунной системе. Ann N Y Acad Sci. 2008, 1143: 170-187. 10.1196 / анналы.1443.017
CAS PubMed Статья Google Scholar
Дэвис В.Л., Хэнкинс М.В., Фостер Р.Г .: Древний опсин и меланопсин позвоночных: расходящиеся детекторы освещенности.Photochem Photobiol Sci. 2010, 9 (11): 1444-1457. 10.1039 / c0pp00203h
CAS PubMed Статья Google Scholar
Wolf G: открытие зрительной функции витамина A. J Nutr. 2001, 131 (6): 1647-1650.
CAS PubMed Google Scholar
Д’Амброзио Д. Н., Клагстон Р. Д., Бланер В. С.: Метаболизм витамина А: обновление. 2011, 3 (1): 63-103.
Penzes P, Napoli JL: Голоклеточный ретинол-связывающий белок: отличие лиганд-связывающего сродства от эффективности в качестве субстрата в биосинтезе сетчатки. Биохимия. 1999, 38 (7): 2088-2093. 10.1021 / bi982228t
КАС PubMed Статья Google Scholar
Лю Л., Гудас Л.Дж .: Нарушение гена лецитин: ретинол-ацилтрансферазы делает мышей более восприимчивыми к дефициту витамина А. J Biol Chem.2005, 280 (48): 40226-40234. 10.1074 / jbc.M509643200
CAS PubMed Статья Google Scholar
Deigner PS, Law WC, Canada FJ, Rando RR: Мембраны как источник энергии в эндергоническом превращении витамина А в 11-цис-ретинол. Наука. 1989, 244 (4907): 968-971. 10.1126 / science.2727688
CAS PubMed Статья Google Scholar
Mata NL, Moghrabi WN, Lee JS, Bui TV, Radu RA, Horwitz J, Travis GH: Rpe65 представляет собой белок, связывающий ретиниловый эфир, который представляет собой нерастворимый субстрат для изомеразы в пигментных эпителиальных клетках сетчатки. J Biol Chem. 2004, 279 (1): 635-643.
CAS PubMed Статья Google Scholar
Саари Дж. К., Наврот М., Кеннеди Б. Н., Гарвин Г. Г., Херли Дж. Б., Хуанг Дж., Поссин Д. Е., Крабб Дж. У.: Нарушение цикла зрения у мышей с нокаутом клеточного ретинальдегид-связывающего белка (CRALBP) приводит к замедленной адаптации к темноте.Нейрон. 2001, 29 (3): 739-748. 10.1016 / S0896-6273 (01) 00248-3
КАС PubMed Статья Google Scholar
Driessen CA, Janssen BP, Winkens HJ, van Vugt AH, de Leeuw TL, Janssen JJ: Клонирование и экспрессия кДНК, кодирующей 11-цис-ретинолдегидрогеназу пигментного пигмента бычьей сетчатки глаза. Инвестируйте Ophthalmol Vis Sci. 1995, 36 (10): 1988–1996.
CAS PubMed Google Scholar
Simon A, Hellman U, Wernstedt C, Eriksson U: 11-цис-ретинолдегидрогеназа, специфичная для пигментного эпителия сетчатки, принадлежит к семейству короткоцепочечных алкогольдегидрогеназ. J Biol Chem. 1995, 270 (3): 1107-1112. 10.1074 / jbc.270.3.1107
КАС PubMed Статья Google Scholar
Duester G: Семейства ретиноиддегидрогеназ, регулирующие функцию витамина А: производство зрительного пигмента и ретиноевой кислоты. Eur J Biochem.2000, 267 (14): 4315-4324. 10.1046 / j.1432-1327.2000.01497.x
CAS PubMed Статья Google Scholar
Агадир А., Нау Х., Бланер В.С.: Ретиноиды: биохимические и молекулярные основы действия витамина А и ретиноидов. 1999 г., Берлин; Нью-Йорк Спрингер.
Google Scholar
Posch KC, Enright WJ, Napoli JL: Синтез ретиноевой кислоты цитозолем из оленьих мышей, отрицательных по алкогольдегидрогеназе.Arch Biochem Biophys. 1989, 274 (1): 171-178. 10.1016 / 0003-9861 (89) -1
КАС PubMed Статья Google Scholar
Ной N: Ретиноид-связывающие белки: медиаторы действия ретиноидов. Biochem J. 2000, 348 (Pt 3): 481-495.
PubMed Central CAS PubMed Статья Google Scholar
Boerman MH, Napoli JL: Характеристика микросомальной ретинолдегидрогеназы: короткоцепочечной алкогольдегидрогеназы с интегральными и периферическими мембранными формами, которая взаимодействует с голо-CRBP (тип I).Биохимия. 1995, 34 (21): 7027-7037. 10.1021 / bi00021a014
CAS PubMed Статья Google Scholar
Лапшина Е.А., Беляева О.В., Чумакова О.В., Кедишвили Н.Ю.: Дифференциальное распознавание свободного и связанного ретинола микросомальными ретинол / стеролдегидрогеназами человека: характеристика активности голо-CRBP-дегидрогеназы RoDH-4. Биохимия. 2003, 42 (3): 776-784. 10.1021 / bi026836r
КАС PubMed Статья Google Scholar
Беляева О.В., Стеценко А.В., Нельсон П., Кедишвили Н.Ю.: Свойства короткоцепочечной дегидрогеназы / редуктазы RalR1: характеристика очищенного фермента, его ориентация в микросомальной мембране и распределение в тканях и линиях клеток человека. Биохимия. 2003, 42 (50): 14838-14845. 10.1021 / bi035288u
CAS PubMed Статья Google Scholar
Парес X, Фаррес Дж., Кедишвили Н., Дестер G: Средне- и короткоцепочечные гены дегидрогеназы / редуктазы и семейства белков: среднецепочечные и короткоцепочечные дегидрогеназы / редуктазы в метаболизме ретиноидов.Cell Mol Life Sci. 2008, 65 (24): 3936-3949. 10.1007 / s00018-008-8591-3
PubMed Central CAS PubMed Статья Google Scholar
Cerignoli F, Guo X, Cardinali B, Rinaldi C, Casaletto J, Frati L, Screpanti I, Gudas LJ, Gulino A, Thiele CJ: retSDR1, короткоцепочечная ретинолдегидрогеназа / редуктаза, представляет собой ретиноевую кислоту -индуцируемые и часто делетированные в клеточных линиях нейробластомы человека. Cancer Res. 2002, 62 (4): 1196-1204.
CAS PubMed Google Scholar
Lei Z, Chen W., Zhang M, Napoli JL: Уменьшение полностью транс-ретиналя в пероксисомной фракции печени мыши с помощью короткоцепочечной дегидрогеназы / редуктазы RRD: индукция клофибратом альфа-лиганда PPAR. Биохимия. 2003, 42 (14): 4190-4196. 10.1021 / bi026948i
КАС PubMed Статья Google Scholar
Crosas B, Hyndman DJ, Gallego O, Martras S, Pares X, Flynn TG, Farres J: Альдозоредуктаза человека и альдозоредуктаза тонкого кишечника человека являются эффективными редуктазами сетчатки: последствия для метаболизма ретиноидов. Biochem J. 2003, 373 (Pt 3): 973-979.
PubMed Central CAS PubMed Статья Google Scholar
Kasus-Jacobi A, Ou J, Bashmakov YK, Shelton JM, Richardson JA, Goldstein JL, Brown MS: Характеристика короткоцепочечной альдегидредуктазы мыши (SCALD), фермента, регулируемого связыванием регуляторных элементов стеролов белки.J Biol Chem. 2003, 278 (34): 32380-32389. 10.1074 / jbc.M304969200
CAS PubMed Статья Google Scholar
Доулинг Дж. Э., Уолд Дж .: Роль витамина А кислоты. Vitam Horm. 1960, 18: 515-541.
CAS PubMed Статья Google Scholar
Duester G, Mic FA, Molotkov A: Цитозольные ретиноиддегидрогеназы управляют повсеместным метаболизмом ретинола в ретинальдегид с последующим тканеспецифическим метаболизмом в ретиноевую кислоту.Chem Biol Interact. 2003, 143-144: 201-210.
CAS PubMed Статья Google Scholar
Haselbeck RJ, Hoffmann I, Duester G: Отдельные функции для Aldh2 и Raldh3 в контроле продукции лиганда для эмбриональных ретиноидных сигнальных путей. Dev Genet. 1999, 25 (4): 353-364. 10.1002 / (SICI) 1520-6408 (1999) 25: 4 <353 :: AID-DVG9> 3.0.CO; 2-G
PubMed Central CAS PubMed Статья Google Scholar
Ang HL, Duester G: фермент биосинтеза ретиноевой кислоты ALDh2 локализуется в подмножестве ретиноид-зависимых тканей во время развития ксенопусов. Dev Dyn. 1999, 215 (3): 264-272. 10.1002 / (SICI) 1097-0177 (199907) 215: 3 <264 :: AID-AJA8> 3.0.CO; 2-I
CAS PubMed Статья Google Scholar
Frota-Ruchon A, Marcinkiewicz M, Bhat PV: Локализация ретинальной дегидрогеназы 1 типа в желудке и кишечнике. Cell Tissue Res.2000, 302 (3): 397-400. 10.1007 / s004410000281
CAS PubMed Статья Google Scholar
Ang HL, Duester G: Стимуляция преждевременного синтеза ретиноевой кислоты у эмбрионов Xenopus после преждевременной экспрессии альдегиддегидрогеназы ALDh2. Eur J Biochem. 1999, 260 (1): 227-234. 10.1046 / j.1432-1327.1999.00139.x
CAS PubMed Статья Google Scholar
Fan X, Molotkov A, Manabe S, Donmoyer CM, Deltour L, Foglio MH, Cuenca AE, Blaner WS, Lipton SA, Duester G: Целевое нарушение Aldh2a1 (Raldh2) свидетельствует о сложном механизме синтеза ретиноевой кислоты в развивающаяся сетчатка. Mol Cell Biol. 2003, 23 (13): 4637-4648. 10.1128 / MCB.23.13.4637-4648.2003
PubMed Central CAS PubMed Статья Google Scholar
Blentic A, Gale E, Maden M: центры передачи сигналов ретиноевой кислоты в эмбрионе птицы, идентифицированные по сайтам экспрессии синтезирующих и катаболизирующих ферментов.Dev Dyn. 2003, 227 (1): 114-127. 10.1002 / dvdy.10292
CAS PubMed Статья Google Scholar
Chen Y, Pollet N, Niehrs C, Pieler T: Повышенная активность XRALDh3 оказывает апостериоризирующее действие на центральную нервную систему эмбрионов Xenopus. Mech Dev. 2001, 101 (1-2): 91-103. 10.1016 / S0925-4773 (00) 00558-X
CAS PubMed Статья Google Scholar
Niederreither K, McCaffery P, Drager UC, Chambon P, Dolle P: Ограниченная экспрессия и индуцированное ретиноевой кислотой подавление гена ретинальдегиддегидрогеназы типа 2 (RALDH-2) во время развития мыши. Mech Dev. 1997, 62 (1): 67-78. 10.1016 / S0925-4773 (96) 00653-3
КАС PubMed Статья Google Scholar
Niederreither K, Subbarayan V, Dolle P, Chambon P: Эмбриональный синтез ретиноевой кислоты необходим для раннего постимплантационного развития мышей.Нат Жене. 1999, 21 (4): 444-448. 10.1038 / 7788
КАС PubMed Статья Google Scholar
Niederreither K, Vermot J, Schuhbaur B, Chambon P, Dolle P: синтез ретиноевой кислоты и формирование паттерна заднего мозга у эмбриона мыши. Разработка. 2000, 127 (1): 75-85.
CAS PubMed Google Scholar
Grandel H, Lun K, Rauch GJ, Rhinn M, Piotrowski T, Houart C, Sordino P, Kuchler AM, Schulte-Merker S, Geisler R: Передача сигналов ретиноевой кислоты у эмбрионов рыбок данио необходима во время пре- этапы сегментации для формирования рисунка передне-задней оси ЦНС и образования зачатка грудного плавника.Разработка. 2002, 129 (12): 2851-2865.
CAS PubMed Google Scholar
Каваками Ю., Райя А., Райя Р.М., Родригес-Эстебан С., Бельмонте Дж. К.: Передача сигналов ретиноевой кислоты связывает лево-правое асимметричное формирование паттерна и билатерально-симметричный сомитогенез в эмбрионе рыбок данио. Природа. 2005, 435 (7039): 165-171. 10.1038 / nature03512
CAS PubMed Статья Google Scholar
Бегеманн Г., Маркс М., Мебус К., Мейер А., Бастмейер М.: Помимо фенотипа шеи: влияние пониженной передачи сигналов ретиноевой кислоты на развитие двигательных нейронов в заднем мозге рыбок данио. Dev Biol. 2004, 271 (1): 119-129. 10.1016 / j.ydbio.2004.03.033
CAS PubMed Статья Google Scholar
Доббс-МакОлифф Б., Чжао К., Линни Э. Механизмы обратной связи регулируют выработку и деградацию ретиноевой кислоты в эмбрионе рыбок данио.Mech Dev. 2004, 121 (4): 339-350. 10.1016 / j.mod.2004.02.008
CAS PubMed Статья Google Scholar
Strate I, Min TH, Iliev D, Pera EM: Ретинолдегидрогеназа 10 является регулятором обратной связи передачи сигналов ретиноевой кислоты во время формирования оси и формирования паттерна центральной нервной системы. Разработка. 2009, 136 (3): 461-472. 10.1242 / dev.024901
CAS PubMed Статья Google Scholar
Koenig SF, Brentle S, Hamdi K, Fichtner D, Wedlich D, Gradl D: En2, Pax2 / 5 и Tcf-4 факторы транскрипции взаимодействуют в формировании паттерна мозга Xenopus. Dev Biol. 340 (2): 318-328.
Lupo G, Liu Y, Qiu R, Chandraratna RA, Barsacchi G, He RQ, Harris WA: формирование дорсовентрального паттерна глаза Xenopus: сотрудничество ретиноидов, Hedgehog и передачи сигналов рецептора FGF. Разработка. 2005, 132 (7): 1737-1748. 10.1242 / dev.01726
CAS PubMed Статья Google Scholar
Mic FA, Molotkov A, Fan X, Cuenca AE, Duester G: RALDh4, ретинальдегиддегидрогеназа, вырабатывающая ретиноевую кислоту, экспрессируется в вентральной части сетчатки, слуховых пузырьках и обонятельной ямке во время развития мышей. Mech Dev. 2000, 97 (1-2): 227-230. 10.1016 / S0925-4773 (00) 00434-2
КАС PubMed Статья Google Scholar
Питтлик С., Домингес С., Мейер А., Бегеманн Г.: Экспрессия рыбок данио aldh2a3 (raldh4) и отсутствие aldh2a1 в костистых костях.Паттерны экспрессии генов. 2008, 8 (3): 141-147. 10.1016 / j.gep.2007.11.003
КАС PubMed Статья Google Scholar
Сузуки Р., Шинтани Т., Сакута Х., Като А., Окавара Т., Осуми Н., Нода М.: Идентификация RALDH-3, новой дегидрогеназы ретинальдегида, экспрессирующейся в вентральной области сетчатки. Mech Dev. 2000, 98 (1-2): 37-50. 10.1016 / S0925-4773 (00) 00450-0
CAS PubMed Статья Google Scholar
Молоткова Н., Молотков А., Дестер Г.: Роль ретиноевой кислоты в развитии переднего мозга начинается поздно, когда Raldh4 генерирует ретиноевую кислоту в вентральной субвентрикулярной зоне. Dev Biol. 2007, 303 (2): 601-610. 10.1016 / j.ydbio.2006.11.035
PubMed Central CAS PubMed Статья Google Scholar
Dupe V, Matt N, Garnier JM, Chambon P, Mark M, Ghyselinck NB: Смертельный дефект новорожденного из-за инактивации ретинальдегиддегидрогеназы 3 типа предотвращается лечением матери ретиноевой кислотой.Proc Natl Acad Sci USA. 2003, 100 (24): 14036-14041. 10.1073 / pnas.2336223100
PubMed Central CAS PubMed Статья Google Scholar
Halilagic A, Ribes V, Ghyselinck NB, Zile MH, Dolle P, Studer M: ретиноиды контролируют передние и дорсальные свойства в развивающемся переднем мозге. Dev Biol. 2007, 303 (1): 362-375. 10.1016 / j.ydbio.2006.11.021
CAS PubMed Статья Google Scholar
Chytil F, Ong DE: Клеточные белки, связывающие ретинол и ретиноевую кислоту в действии витамина А. Fed Proc. 1979, 38 (11): 2510-2514.
CAS PubMed Google Scholar
Napoli JL, Boerman MH, Chai X, Zhai Y, Fiorella PD: ферменты и связывающие белки, влияющие на концентрацию ретиноевой кислоты. J Стероид Biochem Mol Biol. 1995, 53 (1-6): 497-502. 10.1016 / 0960-0760 (95) 00096-I
CAS PubMed Статья Google Scholar
Hoegberg P, Schmidt CK, Fletcher N, Nilsson CB, Trossvik C, Gerlienke Schuur A, Brouwer A, Nau H, Ghyselinck NB, Chambon P: статус ретиноидов и чувствительность к 2, 3, 7, 8-тетрахлордибензо-пара-диоксину (TCDD) у мышей, лишенных ретиноид-связывающего белка или форм ретиноидных рецепторов. Chem Biol Interact. 2005, 156 (1): 25-39. 10.1016 / j.cbi.2005.06.006
CAS PubMed Статья Google Scholar
Sessler RJ, Noy N: сигнал ядерной локализации, активируемый лигандом, в клеточном белке-II, связывающем ретиноевую кислоту.Mol Cell. 2005, 18 (3): 343-353. 10.1016 / j.molcel.2005.03.026
CAS PubMed Статья Google Scholar
Stachurska E, Loboda A, Niderla-Bielinska J, Szperl M, Juszynski M, Jozkowicz A, Dulak J, Ratajska A: Экспрессия клеточного белка, связывающего ретиноевую кислоту I и II (CRABP I и II) в сердца эмбриональных мышей, обработанные ретиноевой кислотой. Acta Biochim Pol. 58 (1): 19-29.
CAS PubMed Google Scholar
Boylan JF, Gudas LJ: Уровень экспрессии CRABP-I влияет на количество и типы метаболитов полностью транс-ретиноевой кислоты в стволовых клетках тератокарциномы F9. J Biol Chem. 1992, 267 (30): 21486-21491.
CAS PubMed Google Scholar
Gu X, Xu F, Wang X, Gao X, Zhao Q: Молекулярное клонирование и экспрессия нового гена CYP26 (cyp26d1) во время раннего развития рыбок данио. Паттерны экспрессии генов. 2005, 5 (6): 733-739.10.1016 / j.modgep.2005.04.005
CAS PubMed Статья Google Scholar
Sakai Y, Luo T, McCaffery P, Hamada H, Drager UC: CYP26A1 и CYP26C1 взаимодействуют в деградации ретиноевой кислоты в экваториальной сетчатке во время более позднего развития глаза. Dev Biol. 2004, 276 (1): 143-157. 10.1016 / j.ydbio.2004.08.032
CAS PubMed Статья Google Scholar
Takeuchi H, Yokota A, Ohoka Y, Iwata M: Cyp26b1 регулирует ретиноевую кислоту-зависимые сигналы в Т-клетках, и его экспрессия подавляется трансформирующим фактором роста-бета. PLoS One. 6 (1): e16089.
Hernandez RE, Putzke AP, Myers JP, Margaretha L, Moens CB: ферменты Cyp26 генерируют паттерн ответа на ретиноевую кислоту, необходимый для развития заднего мозга. Разработка. 2007, 134 (1): 177-187. 10.1242 / dev.02706
PubMed Central CAS PubMed Статья Google Scholar
White JA, Beckett-Jones B, Guo YD, Dilworth FJ, Bonasoro J, Jones G, Petkovich M: клонирование кДНК человеческого фермента, метаболизирующего ретиноевую кислоту (hP450RAI), идентифицирует новое семейство цитохромов P450. J Biol Chem. 1997, 272 (30): 18538-18541. 10.1074 / jbc.272.30.18538
КАС PubMed Статья Google Scholar
de Roos K, Sonneveld E, Compaan B, ten Berge D, Durston AJ, van der Saag PT: Экспрессия 4-гидроксилазы ретиноевой кислоты (CYP26) во время эмбриогенеза мыши и Xenopus laevis.Mech Dev. 1999, 82 (1-2): 205-211. 10.1016 / S0925-4773 (99) 00016-7
CAS PubMed Статья Google Scholar
Swindell EC, Thaller C, Sockanathan S, Petkovich M, Jessell TM, Eichele G: Дополнительные домены продукции и деградации ретиноевой кислоты в раннем курином эмбрионе. Dev Biol. 1999, 216 (1): 282-296. 10.1006 / dbio.1999.9487
CAS PubMed Статья Google Scholar
Giguere V, Ong ES, Segui P, Evans RM: Идентификация рецептора морфогена ретиноевой кислоты. Природа. 1987, 330 (6149): 624-629. 10.1038 / 330624a0
КАС PubMed Статья Google Scholar
Петкович М., Brand NJ, Krust A, Chambon P: рецептор ретиноевой кислоты человека, который принадлежит к семейству ядерных рецепторов. Природа. 1987, 330 (6147): 444-450. 10.1038 / 330444a0
КАС PubMed Статья Google Scholar
Brand N, Petkovich M, Krust A, Chambon P, de The H, Marchio A, Tiollais P, Dejean A: Идентификация второго рецептора ретиноевой кислоты человека. Природа. 1988, 332 (6167): 850-853. 10.1038 / 332850a0
КАС PubMed Статья Google Scholar
Ruberte E, Dolle P, Krust A, Zelent A, Morriss-Kay G, Chambon P: Специфическое пространственное и временное распределение гамма-транскриптов рецепторов ретиноевой кислоты во время эмбриогенеза мышей.Разработка. 1990, 108 (2): 213-222.
CAS PubMed Google Scholar
Li E, Sucov HM, Lee KF, Evans RM, Jaenisch R: Нормальное развитие и рост мышей, несущих нацеленное нарушение гена рецептора альфа-1 ретиноевой кислоты. Proc Natl Acad Sci USA. 1993, 90 (4): 1590-1594. 10.1073 / pnas.90.4.1590
PubMed Central CAS PubMed Статья Google Scholar
Ghyselinck NB, Dupe V, Dierich A, Messaddeq N, Garnier JM, Rochette-Egly C, Chambon P, Mark M: Роль рецептора ретиноевой кислоты бета (RARbeta) во время развития мышей. Int J Dev Biol. 1997, 41 (3): 425-447.
CAS PubMed Google Scholar
Суббараян В., Кастнер П., Марк М., Диерих А., Горри П., Шамбон П. Ограниченная специфичность и большое совпадение функций изоформ RAR гамма 1 и RAR гамма 2 мыши.Mech Dev. 1997, 66 (1-2): 131-142. 10.1016 / S0925-4773 (97) 00098-1
CAS PubMed Статья Google Scholar
Manshouri T, Yang Y, Lin H, Stass SA, Glassman AB, Keating MJ, Albitar M: Подавление RAR-альфа у мышей антисмысловым трансгеном приводит к компенсаторному увеличению RAR-бета и RAR-гамма и развитию лимфома. Кровь. 1997, 89 (7): 2507-2515.
CAS PubMed Google Scholar
Mark M, Ghyselinck NB, Wendling O, Dupe V, Mascrez B, Kastner P, Chambon P: генетическое вскрытие ретиноидного сигнального пути у мышей. Proc Nutr Soc. 1999, 58 (3): 609-613. 10.1017 / S002966519
CAS PubMed Статья Google Scholar
Chomienne C, Balitrand N, Ballerini P, Castaigne S, de The H, Degos L: полностью транс-ретиноевая кислота модулирует рецептор ретиноевой кислоты-альфа в промиелоцитарных клетках.J Clin Invest. 1991, 88 (6): 2150-2154. 10.1172 / JCI115547
PubMed Central CAS PubMed Статья Google Scholar
Kamei Y, Kawada T, Kazuki R, Sugimoto E: Экспрессия гена гамма-2 рецептора ретиноевой кислоты повышается с помощью ретиноевой кислоты в преадипоцитах 3T3-L1. Biochem J. 1993, 293 (Pt 3): 807-812.
PubMed Central CAS PubMed Статья Google Scholar
Sucov HM, Murakami KK, Evans RM: Характеристика ауторегулируемого ответного элемента в гене бета-типа рецептора ретиноевой кислоты мыши. Proc Natl Acad Sci USA. 1990, 87 (14): 5392-5396. 10.1073 / pnas.87.14.5392
PubMed Central CAS PubMed Статья Google Scholar
Чавла А., Репа Дж. Дж., Эванс Р. М., Мангельсдорф Д. Д.: Ядерные рецепторы и физиология липидов: открытие X-файлов. Наука. 2001, 294 (5548): 1866-1870.10.1126 / science.294.5548.1866
CAS PubMed Статья Google Scholar
Kastner P, Messaddeq N, Mark M, Wendling O, Grondona JM, Ward S, Ghyselinck N, Chambon P: Дефицит витамина A и мутации RXRalpha, RXRbeta и RARalpha приводят к ранней дифференцировке кардиомиоцитов эмбрионального желудочка. Разработка. 1997, 124 (23): 4749-4758.
CAS PubMed Google Scholar
Kastner P, Mark M, Leid M, Gansmuller A, Chin W, Grondona JM, Decimo D, Krezel W, Dierich A, Chambon P: аномальный сперматогенез у мышей с мутантом RXR beta. Genes Dev. 1996, 10 (1): 80-92. 10.1101 / gad.10.1.80
CAS PubMed Статья Google Scholar
Krezel W, Dupe V, Mark M, Dierich A, Kastner P, Chambon P: мыши с нулевым гамма-тестом RXR, по-видимому, нормальны, а соединение RXR alpha + / — / RXR beta — / — / RXR гамма — / — мутантные мыши жизнеспособны.Proc Natl Acad Sci USA. 1996, 93 (17): 9010-9014.
PubMed Central CAS PubMed Статья Google Scholar
Mello T, Polvani S, Galli A: Рецептор, активируемый пролифератором пероксисом, и ретиноевый рецептор x при алкогольной болезни печени. PPAR Res. 2009, 2009: 748174.
PubMed Central PubMed Статья CAS Google Scholar
Rizzo G, Renga B, Antonelli E, Passeri D, Pellicciari R, Fiorucci S: Метилтрансфераза PRMT1 действует как коактиватор фарнезоидного рецептора X (FXR) / 9-цис-ретиноидного рецептора X и регулирует транскрипцию FXR-чувствительных генов.Mol Pharmacol. 2005, 68 (2): 551-558. 10.1124 / моль. 105.012104
CAS PubMed Статья Google Scholar
Wingert RA, Davidson AJ: Pronephros рыбок данио: модель для изучения сегментации нефронов. Kidney Int. 2008, 73 (10): 1120-1127. 10.1038 / ки.2008.37
КАС PubMed Статья Google Scholar
Lagu B, Pio B, Lebedev R, Yang M, Pelton PD: RXR-LXR гетеродимерные модуляторы для потенциального лечения дислипидемии.Bioorg Med Chem Lett. 2007, 17 (12): 3497-3503. 10.1016 / j.bmcl.2007.01.047
CAS PubMed Статья Google Scholar
Чжао С., Дальман-Райт К.: Х-рецептор печени в метаболизме холестерина. J Endocrinol. 204 (3): 233-240.
Desvergne B: RXR: от партнерства к лидерству в регуляции метаболизма. Vitam Horm. 2007, 75: 1-32.
CAS PubMed Статья Google Scholar
Desvergne B, Wahli W: Рецепторы, активируемые пролифератором пероксисом: ядерный контроль метаболизма. Endocr Rev.1999, 20 (5): 649-688. 10.1210 / er.20.5.649
CAS PubMed Google Scholar
Desvergne B, Michalik L, Wahli W: Транскрипционная регуляция метаболизма. Physiol Rev.2006, 86 (2): 465-514. 10.1152 / Physrev.00025.2005
КАС PubMed Статья Google Scholar
Hollemann T, Chen Y, Grunz H, Pieler T: Региональная метаболическая активность устанавливает границы передачи сигналов ретиноевой кислоты. EMBO J. 1998, 17 (24): 7361-7372. 10.1093 / emboj / 17.24.7361
PubMed Central CAS PubMed Статья Google Scholar
Astrom A, Pettersson U, Voorhees JJ: Структура гена человеческого клеточного белка II, связывающего ретиноевую кислоту. Ранняя регуляция транскрипции ретиноевой кислотой. J Biol Chem.1992, 267 (35): 25251-25255.
CAS PubMed Google Scholar
Durand B, Saunders M, Leroy P, Leid M, Chambon P: индукция транскрипции CRABPII полностью транс- и 9-цис ретиноевой кислотой опосредуется гетеродимерами RAR-RXR, связанными с повторяющимися мотивами DR1 и DR2. Клетка. 1992, 71 (1): 73-85. 10.1016 / 0092-8674 (92)
-GCAS PubMed Статья Google Scholar
Zhu J, Heyworth CM, Glasow A, Huang QH, Petrie K, Lanotte M, Benoit G, Gallagher R, Waxman S, Enver T: Линейное ограничение экспрессии гена RARalpha в миелоидной дифференцировке. Кровь. 2001, 98 (8): 2563-2567. 10.1182 / blood.V98.8.2563
CAS PubMed Статья Google Scholar
Takeyama K, Kojima R, Ohashi R, Sato T, Mano H, Masushige S, Kato S. Ретиноевая кислота по-разному регулирует экспрессию гена альфа- и гамма-изоформ рецептора ретиноевой кислоты у эмбрионов и взрослых крыс.Biochem Biophys Res Commun. 1996, 222 (2): 395-400. 10.1006 / bbrc.1996.0755
CAS PubMed Статья Google Scholar
Halme A, Cheng M, Hariharan IK: Ретиноиды регулируют контрольную точку развития для регенерации тканей у Drosophila. Curr Biol. 20 (5): 458-463.
Маден М: Ретиноиды в эмбрионах не млекопитающих. Методы Мол биол. 2008, 461: 541-559. 10.1007 / 978-1-60327-483-8_37
CAS PubMed Статья Google Scholar
Uehara M, Yashiro K, Takaoka K, Yamamoto M, Hamada H: Удаление материнской ретиноевой кислоты эмбриональным CYP26 необходимо для правильной экспрессии Nodal во время раннего формирования эмбрионального паттерна. Genes Dev. 2009, 23 (14): 1689-1698. 10.1101 / gad.1776209
PubMed Central CAS PubMed Статья Google Scholar
Кудох Т., Уилсон С.В., Давид И.Б.: Определенные роли Fgf, Wnt и ретиноевой кислоты в постериоризации нервной эктодермы.Разработка. 2002, 129 (18): 4335-4346.
CAS PubMed Google Scholar
White RJ, Schilling TF: Как деградирует: Cyp26s в развитии заднего мозга. Dev Dyn. 2008, 237 (10): 2775-2790. 10.1002 / dvdy.21695
PubMed Central CAS PubMed Статья Google Scholar
Kessel M, Gruss P: Гомеотические трансформации позвонков мыши и сопутствующее изменение Hox-кодов, вызванное ретиноевой кислотой.Клетка. 1991, 67 (1): 89-104. 10.1016 / 0092-8674 (91) -I
CAS PubMed Статья Google Scholar
Padmanabhan R: индуцированный ретиноевой кислотой синдром каудальной регрессии у плода мыши. Reprod Toxicol. 1998, 12 (2): 139-151. 10.1016 / S0890-6238 (97) 00153-6
КАС PubMed Статья Google Scholar
Durston AJ, Timmermans JP, Hage WJ, Hendriks HF, de Vries NJ, Heideveld M, Nieuwkoop PD: Ретиноевая кислота вызывает переднезаднюю трансформацию в развивающейся центральной нервной системе.Природа. 1989, 340 (6229): 140-144. 10.1038 / 340140a0
КАС PubMed Статья Google Scholar
Sive HL, Draper BW, Harland RM, Weintraub H: Идентификация периода, чувствительного к ретиноевой кислоте, во время формирования первичной оси у Xenopus laevis. Genes Dev. 1990, 4 (6): 932-942. 10.1101 / gad.4.6.932
CAS PubMed Статья Google Scholar
Ruiz i Altaba A, Jessell TM: Ретиноевая кислота изменяет структуру дифференцировки клеток в центральной нервной системе у эмбрионов Xenopus на стадии нейрулы. Разработка. 1991, 112 (4): 945-958.
CAS PubMed Google Scholar
Chazaud C, Chambon P, Dolle P: Ретиноевая кислота необходима эмбриону мыши для определения лево-правой асимметрии и морфогенеза сердца. Разработка. 1999, 126 (12): 2589-2596.
CAS PubMed Google Scholar
Huang S, Ma J, Liu X, Zhang Y, Luo L: передача сигналов ретиноевой кислоты последовательно контролирует латеральность внутренних органов и сердца у рыбок данио. J Biol Chem. 286 (32): 28533-28543.
Вермот Дж., Галлего Ллама Дж., Фраулоб В., Нидеррайтер К., Шамбон П., Долле П. Ретиноевая кислота контролирует двустороннюю симметрию образования сомитов в эмбрионе мыши. Наука. 2005, 308 (5721): 563-566. 10.1126 / science.1108363
CAS PubMed Статья Google Scholar
Vermot J, Pourquie O: Ретиноевая кислота координирует сомитогенез и формирование левого-правого паттерна у эмбрионов позвоночных. Природа. 2005, 435 (7039): 215-220. 10.1038 / nature03488
CAS PubMed Статья Google Scholar
Anderson DJ: Происхождение и факторы транскрипции в спецификации первичных сенсорных нейронов позвоночных. Curr Opin Neurobiol. 1999, 9 (5): 517-524. 10.1016 / S0959-4388 (99) 00015-X
CAS PubMed Статья Google Scholar
Ribes V, Stutzmann F, Bianchetti L, Guillemot F, Dolle P, Le Roux I: Комбинаторная передача сигналов контролирует экспрессию Neurogenin2 в начале спинального нейрогенеза. Dev Biol. 2008, 321 (2): 470-481. 10.1016 / j.ydbio.2008.06.003
CAS PubMed Статья Google Scholar
Coyle DE, Li J, Baccei M: региональная дифференцировка нейронов стволовых клеток плюрипотентной эмбриональной карциномы человека, индуцированных ретиноевой кислотой. PLoS One.2011, 6 (1): e16174. 10.1371 / journal.pone.0016174
PubMed Central CAS PubMed Статья Google Scholar
Shan ZY, Liu F, Lei L, Li QM, Jin LH, Wu YS, Li X, Shen JL: Создание ГАМКергических нейронов спинного мозга из эмбриональных стволовых клеток мыши. Перепрограммирование клеток. 2011, 13 (1): 85-91.
CAS PubMed Статья Google Scholar
Chen N, Napoli JL: Полностью транс-ретиноевая кислота стимулирует трансляцию и индуцирует образование шипов в нейронах гиппокампа через ассоциированный с мембраной RARalpha. FASEB J. 2008, 22 (1): 236-245.
CAS PubMed Статья Google Scholar
Krumlauf R: Hox-гены и формирование паттерна в жаберной области головы позвоночных. Тенденции Genet. 1993, 9 (4): 106-112. 10.1016 / 0168-9525 (93)
КАС PubMed Статья Google Scholar
Studer M, Lumsden A, Ariza-McNaughton L, Bradley A, Krumlauf R: измененная сегментарная идентичность и аномальная миграция моторных нейронов у мышей, лишенных Hoxb-1. Природа. 1996, 384 (6610): 630-634. 10.1038 / 384630a0
КАС PubMed Статья Google Scholar
Marshall H, Nonchev S, Sham MH, Muchamore I., Lumsden A, Krumlauf R: Ретиноевая кислота изменяет Hox-код заднего мозга и индуцирует преобразование ромбомеров 2/3 в идентичность 4/5.Природа. 1992, 360 (6406): 737-741. 10.1038 / 360737a0
CAS PubMed Статья Google Scholar
Kessel M: Обращение аксональных путей от ромбомера 3 коррелирует с дополнительными доменами экспрессии Hox. Нейрон. 1993, 10 (3): 379-393. 10.1016 / 0896-6273 (93) -O
CAS PubMed Статья Google Scholar
Hill J, Clarke JD, Vargesson N, Jowett T., Holder N: Экзогенная ретиноевая кислота вызывает специфические изменения в развитии среднего и заднего мозга эмбриона рыбок данио, включая позиционную повторную спецификацию маутнеровского нейрона.Mech Dev. 1995, 50 (1): 3-16. 10.1016 / 0925-4773 (94) 00321-D
CAS PubMed Статья Google Scholar
Hernandez RE, Rikhof HA, Bachmann R, Moens CB: vhnf1 интегрирует глобальное формирование паттерна RA и локальные сигналы FGF, чтобы направлять развитие заднего заднего мозга у рыбок данио. Разработка. 2004, 131 (18): 4511-4520. 10.1242 / dev.01297
CAS PubMed Статья Google Scholar
Wiellette EL, Sive H: vhnf1 и Fgf сигналы синергизируют, чтобы специфицировать идентичность ромбомеров в заднем мозге рыбок данио. Разработка. 2003, 130 (16): 3821-3829. 10.1242 / dev.00572
CAS PubMed Статья Google Scholar
Begemann G, Meyer A: Повторное рассмотрение паттерна заднего мозга: время и эффекты передачи сигналов ретиноевой кислоты. Биологические исследования. 2001, 23 (11): 981-986. 10.1002 / bies.1142
CAS PubMed Статья Google Scholar
Гавалас A: Организация заднего мозга. Trends Neurosci. 2002, 25 (2): 61-64. 10.1016 / S0166-2236 (02) 02067-2
КАС PubMed Статья Google Scholar
Dupe V, Lumsden A: Формирование паттерна заднего мозга включает дифференцированные ответы на передачу сигналов ретиноевой кислоты. Разработка. 2001, 128 (12): 2199-2208.
CAS PubMed Google Scholar
Begemann G, Schilling TF, Rauch GJ, Geisler R, Ingham PW: Мутация без шеи у рыбок данио выявляет потребность в raldh3 в мезодермальных сигналах, которые формируют структуру заднего мозга.Разработка. 2001, 128 (16): 3081-3094.
CAS PubMed Google Scholar
Partanen J: Пути передачи сигналов FGF в развитии среднего и переднего мозга. J Neurochem. 2007, 101 (5): 1185-1193. 10.1111 / j.1471-4159.2007.04463.x
CAS PubMed Статья Google Scholar
Emoto Y, Wada H, Okamoto H, Kudo A, Imai Y: Фермент Cyp26a1, метаболизирующий ретиноевую кислоту, необходим для определения территорий заднего и спинного мозга у рыбок данио.Dev Biol. 2005, 278 (2): 415-427. 10.1016 / j.ydbio.2004.11.023
CAS PubMed Статья Google Scholar
Chen Y, Pan FC, Brandes N, Afelik S, Solter M, Pieler T: Передача сигналов ретиноевой кислоты необходима для развития поджелудочной железы и способствует эндокринной системе за счет дифференцировки экзокринных клеток у Xenopus. Dev Biol. 2004, 271 (1): 144-160. 10.1016 / j.ydbio.2004.03.030
CAS PubMed Статья Google Scholar
Stafford D, Hornbruch A, Mueller PR, Prince VE: консервативная роль ретиноидной передачи сигналов в развитии поджелудочной железы позвоночных. Dev Genes Evol. 2004, 214 (9): 432-441.
CAS PubMed Статья Google Scholar
Martin M, Gallego-Llamas J, Ribes V, Kedinger M, Niederreither K, Chambon P, Dolle P, Gradwohl G: Агенез дорсальной поджелудочной железы у мутантных мышей Raldh3 с дефицитом ретиноевой кислоты. Dev Biol. 2005, 284 (2): 399-411.10.1016 / j.ydbio.2005.05.035
CAS PubMed Статья Google Scholar
Molotkov A, Molotkova N, Duester G: Ретиноевая кислота, вырабатываемая Raldh3 в мезодерме, необходима для развития дорсальной энтодермальной поджелудочной железы мыши. Dev Dyn. 2005, 232 (4): 950-957. 10.1002 / dvdy.20256
CAS PubMed Статья Google Scholar
Pan FC, Chen Y, Bayha E, Pieler T: Опосредованное ретиноевой кислотой формирование паттерна препанкреатической энтодермы у Xenopus действует через прямые и косвенные механизмы.Mech Dev. 2007, 124 (7-8): 518-531. 10.1016 / j.mod.2007.06.003
CAS PubMed Статья Google Scholar
Bayha E, Jorgensen MC, Serup P, Grapin-Botton A: Передача сигналов ретиноевой кислоты организует спецификацию энтодермального органа вдоль всей переднезадней оси. PLoS One. 2009, 4 (6): e5845. 10.1371 / journal.pone.0005845
PubMed Central PubMed Статья CAS Google Scholar
Stafford D, Prince VE: Передача сигналов ретиноевой кислоты необходима для критического раннего этапа развития поджелудочной железы у рыбок данио. Curr Biol. 2002, 12 (14): 1215-1220. 10.1016 / S0960-9822 (02) 00929-6
КАС PubMed Статья Google Scholar
Stafford D, White RJ, Kinkel MD, Linville A, Schilling TF, Prince VE: ретиноиды передают сигнал непосредственно энтодерме рыбок данио для определения экспрессирующих инсулин бета-клеток. Разработка. 2006, 133 (5): 949-956.10.1242 / dev.02263
CAS PubMed Статья Google Scholar
Dalgin G, Ward AB, Hao le T, Beattie CE, Nechiporuk A, Prince VE: mnx1 рыбок данио контролирует выбор судьбы клеток в развивающейся эндокринной поджелудочной железе. Разработка. 138 (21): 4597-4608.
Кинкель, доктор медицины, принц В.Э .: В меню диабетиков: рыбки данио как модель развития и функционирования поджелудочной железы. Биологические исследования. 2009, 31 (2): 139-152. 10.1002 / bies.200800123
PubMed Central CAS PubMed Статья Google Scholar
Kinkel MD, Eames SC, Alonzo MR, Prince VE: Cdx4 необходим в энтодерме для локализации поджелудочной железы и ограничения количества бета-клеток. Разработка. 2008, 135 (5): 919-929. 10.1242 / dev.010660
CAS PubMed Статья Google Scholar
Ostrom M, Loffler KA, Edfalk S, Selander L, Dahl U, Ricordi C, Jeon J, Correa-Medina M, Diez J, Edlund H: Ретиноевая кислота способствует образованию предшественников эндокринных клеток поджелудочной железы и их дальнейшая дифференцировка в бета-клетки.PLoS One. 2008, 3 (7): e2841. 10.1371 / journal.pone.0002841
PubMed Central PubMed Статья CAS Google Scholar
Тулачан С.С., Дои Р., Кавагути Ю., Цудзи С., Накадзима С., Масуи Т., Коидзуми М., Тойода Е., Мори Т., Ито D: полностью транс-ретиноевая кислота индуцирует дифференцировку протоков и эндокринных клеток мезенхимальными / эпителиальные взаимодействия в поджелудочной железе эмбриона. Сахарный диабет. 2003, 52 (1): 76-84. 10,2337 / диабет 52.1,76
КАС PubMed Статья Google Scholar
Шен С.Н., Маргари А., Чиен С.Й., Диксон С., Слэк Дж. М., Тош Д. Полностью транс-ретиноевая кислота подавляет экзокринную дифференцировку и морфогенез ветвления в поджелудочной железе эмбриона. Дифференциация. 2007, 75 (1): 62-74.
PubMed Central CAS PubMed Статья Google Scholar
Мория Н., Комазаки С., Такахаши С., Йокота С., Асашима М: образование поджелудочной железы in vitro из эктодермы Xenopus, обработанной активином и ретиноевой кислотой.Dev Growth Differ. 2000, 42 (6): 593-602. 10.1046 / j.1440-169x.2000.00542.x
CAS PubMed Статья Google Scholar
Asashima M, Michiue T, Kurisaki A: Выяснение роли активина в органогенезе с использованием системы индукции множества органов с недифференцированными клетками амфибий и мышей in vitro. Dev Growth Differ. 2008, 50 (Приложение 1): S35-45.
CAS PubMed Статья Google Scholar
Heine UI, Roberts AB, Munoz EF, Roche NS, Sporn MB: Влияние дефицита ретиноидов на развитие сердца и сосудистой системы эмбриона перепела. Вирхова Arch B Cell Pathol Incl Mol Pathol. 1985, 50 (2): 135-152.
CAS PubMed Google Scholar
Lin SC, Dolle P, Ryckebusch L, Noseda M, Zaffran S, Schneider MD, Niederreither K: Эндогенная ретиноевая кислота регулирует дифференцировку сердечных предшественников.Proc Natl Acad Sci USA. 2010, 107 (20): 9234-9239. 10.1073 / pnas.0
0107PubMed Central CAS PubMed Статья Google Scholar
Sirbu IO, Zhao X, Duester G: Ретиноевая кислота контролирует формирование переднезаднего паттерна сердца путем подавления Isl1 через путь Fgf8. Dev Dyn. 2008, 237 (6): 1627-1635. 10.1002 / dvdy.21570
PubMed Central CAS PubMed Статья Google Scholar
Stachurska E, Loboda A, Niderla-Bielinska J, Szperl M, Juszynski M, Jozkowicz A, Dulak J, Ratajska A: Экспрессия клеточного белка, связывающего ретиноевую кислоту I и II (CRABP I и II) в сердцах эмбриональных мышей, обработанных ретиноевая кислота. Acta Biochim Pol. 2011, 58 (1): 19-29.
CAS PubMed Google Scholar
Keegan BR, Feldman JL, Begemann G, Ingham PW, Yelon D: Передача сигналов ретиноевой кислоты ограничивает пул сердечных предшественников.Наука. 2005, 307 (5707): 247-249. 10.1126 / science.1101573
CAS PubMed Статья Google Scholar
Ryckebusch L, Wang Z, Bertrand N, Lin SC, Chi X, Schwartz R, Zaffran S, Niederreither K: Дефицит ретиноевой кислоты изменяет формирование второго поля сердца. Proc Natl Acad Sci USA. 2008, 105 (8): 2913-2918. 10.1073 / pnas.0712344105
PubMed Central CAS PubMed Статья Google Scholar
Mathew LK, Sengupta S, Franzosa JA, Perry J, La Du J, Andreasen EA, Tanguay RL: Сравнительное профилирование экспрессии показывает важную роль raldh3 в эпиморфной регенерации. J Biol Chem. 2009, 284 (48): 33642-33653. 10.1074 / jbc.M109.011668
PubMed Central CAS PubMed Статья Google Scholar
Кикучи К., Холдуэй Дж. Э., Майор Р. Дж., Блюм Н., Дан Р. Д., Бегеманн Дж., Посс К. Д. Производство ретиноевой кислоты эндокардом и эпикардием является ответной реакцией на травмы, необходимой для регенерации сердца рыбок данио.Dev Cell. 20 (3): 397-404.
Serluca FC, Fishman MC: Предварительный паттерн в пронефрическом поле почек рыбок данио. Разработка. 2001, 128 (12): 2233-2241.
CAS PubMed Google Scholar
Cartry J, Nichane M, Ribes V, Colas A, Riou JF, Pieler T, Dolle P, Bellefroid EJ, Umbhauer M: передача сигналов ретиноевой кислоты необходима для спецификации судьбы пронефрических клеток. Dev Biol. 2006, 299 (1): 35-51.10.1016 / j.ydbio.2006.06.047
CAS PubMed Статья Google Scholar
Мендельсон С., Лонес Д., Децимо Д., Лафкин Т., Лемер М., Шамбон П., Марк М.: Функция рецепторов ретиноевой кислоты (RAR) во время развития (II). Множественные аномалии на разных этапах органогенеза у двойных мутантов RAR. Разработка. 1994, 120 (10): 2749-2771.
CAS PubMed Google Scholar
Carroll TJ, Vize PD: синергизм между Pax-8 и lim-1 в эмбриональном развитии почек. Dev Biol. 1999, 214 (1): 46-59. 10.1006 / dbio.1999.9414
CAS PubMed Статья Google Scholar
Lee SJ, Kim S, Choi SC, Han JK: XPteg (ген, экспрессируемый в проксимальных канальцах Xenopus) важен для спецификации пронефрической мезодермы и тубулогенеза. Mech Dev. 2010, 127 (1-2): 49-61. 10.1016 / j.mod.2009.11.001
CAS PubMed Статья Google Scholar
Bollig F, Perner B, Besenbeck B, Kothe S, Ebert C, Taudien S, Englert C: высококонсервативный элемент, чувствительный к ретиноевой кислоте, контролирует экспрессию wt1a в пронефросе рыбок данио. Разработка. 2009, 136 (17): 2883-2892. 10.1242 / dev.031773
CAS PubMed Статья Google Scholar
Пернер Б., Энглерт С., Боллиг Ф .: Гены опухоли Вильмса wt1a и wt1b контролируют различные стадии во время формирования пронефроса рыбок данио.Dev Biol. 2007, 309 (1): 87-96. 10.1016 / j.ydbio.2007.06.022
CAS PubMed Статья Google Scholar
Osafune K, Nishinakamura R, Komazaki S, Asashima M: индукция пронефрического протока in vitro в эксплантатах Xenopus. Dev Growth Differ. 2002, 44 (2): 161-167. 10.1046 / j.1440-169x.2002.00631.x
PubMed Статья Google Scholar
Wingert RA, Selleck R, Yu J, Song HD, Chen Z, Song A, Zhou Y, Thisse B, Thisse C, McMahon AP: гены cdx и ретиноевая кислота контролируют позиционирование и сегментацию рыбок данио пронефрос.PLoS Genet. 2007, 3 (10): 1922-1938.
CAS PubMed Статья Google Scholar
Росселот С., Спраггон Л., Чиа I, Батурина Е., Риччио П., Лу Б., Нидеррайтер К., Долле П., Дестер Г., Шамбон П. Не клеточно-автономная ретиноидная передача сигналов имеет решающее значение для развития почек. Разработка. 137 (2): 283-292.
Desai TJ, Malpel S, Flentke GR, Smith SM, Cardoso WV: Ретиноевая кислота избирательно регулирует экспрессию Fgf10 и поддерживает клеточную идентичность в предполагаемом поле легких развивающейся передней кишки.Dev Biol. 2004, 273 (2): 402-415. 10.1016 / j.ydbio.2004.04.039
CAS PubMed Статья Google Scholar
Desai TJ, Chen F, Lu J, Qian J, Niederreither K, Dolle P, Chambon P, Cardoso WV: отчетливые роли альфа- и бета-рецепторов ретиноевой кислоты в раннем морфогенезе легких. Dev Biol. 2006, 291 (1): 12-24. 10.1016 / j.ydbio.2005.10.045
CAS PubMed Статья Google Scholar
Chen F, Cao Y, Qian J, Shao F, Niederreither K, Cardoso WV: Зависящая от ретиноевой кислоты сеть в передней кишке контролирует формирование зачатка легкого мыши. J Clin Invest. 120 (6): 2040-2048.
Мальпель С., Мендельсон С., Кардосо В.В.: Регулирование передачи сигналов ретиноевой кислоты во время морфогенеза легких. Разработка. 2000, 127 (14): 3057-3067.
CAS PubMed Google Scholar
Shiotsugu J, Katsuyama Y, Arima K, Baxter A, Koide T, Song J, Chandraratna RA, Blumberg B: множественные точки взаимодействия между ретиноевой кислотой и передачей сигналов FGF во время формирования эмбриональной оси.Разработка. 2004, 131 (11): 2653-2667. 10.1242 / dev.01129
CAS PubMed Статья Google Scholar
Ho L, Mercola M, Gudas LJ: Xenopus laevis клеточный белок, связывающий ретиноевую кислоту: временной и пространственный паттерн экспрессии во время раннего эмбриогенеза. Mech Dev. 1994, 47 (1): 53-64. 10.1016 / 0925-4773 (94)-7
КАС PubMed Статья Google Scholar
Kam RK, Chen Y, Chan SO, Chan WY, Dawid IB, Zhao H: Экспрессия развития короткоцепочечной дегидрогеназы / редуктазы Xenopus 3. Int J Dev Biol. 2010, 54 (8-9): 1355-1360. 10.1387 / ijdb.0
rk
PubMed Central CAS PubMed Статья Google Scholar
Обнаружение межвиткового короткого замыкания статора асинхронного двигателя в соответствии с компонентами последовательности тока линии с использованием искусственной нейронной сети
Обнаружение неисправности предназначено для обнаружения неисправности на начальной стадии и немедленного отключения машины, чтобы избежать отказа двигателя из-за большой ток короткого замыкания.В этой работе представлена онлайн-диагностика межвиткового замыкания статора трехфазного асинхронного двигателя на основе концепции симметричных компонентов. Математическая модель асинхронного двигателя с повреждением при повороте разработана для интерпретации характеристик машины при неисправности. Модель Simulink трехфазного асинхронного двигателя с межвитковым замыканием статора создана для выделения составляющих последовательности тока и напряжения. Ток обратной последовательности может обеспечить решающий и быстрый метод контроля для обнаружения межвиткового короткого замыкания статора асинхронного двигателя.Изменение на единицу тока обратной последовательности по отношению к току прямой последовательности является основным индикатором неисправности, который импортируется в архитектуру нейронной сети. Выходной сигнал нейронной сети прямого обратного распространения классифицирует уровень короткого замыкания обмотки статора.
1. Введение
Асинхронные двигатели превосходят область электромеханического преобразования энергии. Их надежность, низкая стоимость и высокая производительность делают их самыми популярными двигателями переменного тока.Эти двигатели могут применяться в различных областях, от бытовых приборов до промышленных двигателей большой мощности. В последние годы проблемы отказов в больших асинхронных двигателях стали более значительными. Для диагностики неисправностей важно определить, есть ли в системе неисправность, и найти ее источник [1]. Если неисправность двигателя не будет устранена на ранней стадии, это может привести к повреждению двигателя. Это вызовет остановку промышленного производства.
В [2] упоминается много аварийных ситуаций.Одним из них является случай, когда разрушенная штанга ротора вырвалась из паза и повредила обмотку статора. Неисправности асинхронного двигателя могут быть механическими или электрическими. Основные механические неисправности — это неисправность подшипника [3–5] и поломка стержня ротора [6–10]. На электрическую неисправность влияет качество электроэнергии, подаваемой в сеть переменного тока, колебания частоты, нарушения напряжения и колебания нагрузки. Еще одна неисправность — короткое замыкание обмотки статора [3, 11–14]. Примерно более одной трети всех неисправностей, возникающих в асинхронном двигателе, приходится на неисправность обмотки статора.Короткое замыкание в обмотке статора развивается за очень короткое время и полностью выходит из строя. Обычно межвитковое короткое замыкание перерастает в межобмоточное замыкание, замыкание фазной обмотки и замыкание на землю одной линии, что приводит к поломке двигателя. Обнаружение неисправности обмотки на этапе пуска увеличивает возможность ремонта машины путем ее перемотки или, в больших двигателях, смещения короткозамкнутых катушек.
Традиционные способы контроля неисправностей касались измерения потока утечки [15], частичного разряда [16], гармоник в токе и напряжении статора [17] и т. Д.Последующие исследования, однако, показали, что многие из этих традиционных методов могут быть необычными из-за искажений напряжения питания [18], встроенной асимметрии машины [19], случайных последствий отказов статора и ротора и т. Д. Анализ сигнатуры тока двигателя (MCSA). ) является важным методом, принятым для мониторинга состояния. Неисправности асинхронного двигателя, такие как проблемы с подшипниками, поломка стержня ротора, аномалии эксцентриситета и неисправности обмотки статора, вызывают изменение амплитуды и частоты сигнатуры тока двигателя [3–9, 11–14].
Прорыв в технологиях формирования сигналов и достижения в области компьютерного программного обеспечения подняли обнаружение неисправностей машин на новый уровень. Большая часть продемонстрированной работы по обнаружению неисправности обмотки статора относится к области частотного анализа. Методы преобразования сигналов, такие как быстрое преобразование Фурье (FFT), S-преобразование, кратковременное преобразование Фурье (STFT), вейвлет-преобразование и преобразования Гильберта, были приняты в сочетании с различными методами классификации, такими как экспертные системы, искусственная нейронная сеть, нечеткая логика. , и поддерживающая векторная машина [20–26] для моторной деградации.
В [27–29] большой интерес был проявлен к искусственной нейронной сети для обнаружения неисправностей асинхронного двигателя. Необходимым условием для создания успешного классификатора ИНС является выбор соответствующих входных данных для каждого случая неисправности. В [27, 28] описывается обнаружение местоположения межвиткового замыкания статора посредством ИНС с учетом параметров частотной области как выбранного входа.
В [29] ИНС применяется для обнаружения серьезности межобмоточного замыкания с выбранным параметром во временной области.В работах [30–33] основное внимание уделяется току обратной последовательности, который возникает из-за несимметричных обмоток.
Большая часть исследовательских работ по обнаружению неисправности обмотки статора асинхронного двигателя основана на анализе частотной области. Если мы выберем линейные токи или линейные напряжения в качестве параметров, рассматриваемых для обнаружения неисправности, анализ во временной области также будет столь же эффективным. Это позволит избежать использования спектрального анализатора и сложных методов преобразования сигнала, что значительно упрощает системный блок обнаружения неисправностей.Цель состоит в том, чтобы определить универсальный метод диагностики для обнаружения неисправности обмотки и уровня ее серьезности без данных о конструкции двигателя и со знанием параметров неисправности из анализа во временной области.
В этой работе мы пытаемся найти метод обнаружения повреждения обмотки статора на основе удельного значения составляющих последовательности тока во временной области и классифицировать серьезность повреждения с помощью искусственной нейронной сети. Здесь сеть была обучена с полным диапазоном входных векторов, полученных из модели Simulink.Входной вектор NN содержит экспериментальные значения до возможного диапазона и значения Simulink для завершения входного набора. Это обеспечивает хорошо обученную сеть. Значения моделирования хорошо согласуются с экспериментальными значениями. Обнаружение неисправностей на начальном этапе увеличивает жизнеспособность ремонта машины, а устранение неисправностей в зародыше позволяет избежать электрических искр и взрывов.
Следующая сессия подробно описывает математическое моделирование асинхронного двигателя с повреждением обмотки статора. Используя эти математические уравнения, в Simulink создается модель, которая описана в Разделе 3.Метод классификации, используемый для обнаружения неисправности, подчеркивается в разделе 4.
2. Математическое моделирование неисправности витка обмотки статора
Рассматривается трехфазный асинхронный двигатель с повреждением однофазной обмотки статора, где β — это доля закороченных витков. Обмотка в этой фазе состоит из двух частей — витков с коротким замыканием и витков без повреждения. Машинные уравнения в переменных abc для симметричного двигателя с повреждением одной обмотки могут быть выражены как [34–36].Здесь мы предположили, что индуктивность рассеяния закороченных витков равна, где — индуктивность рассеяния по фазе, а полное сопротивление короткого замыкания — резистивное. Где
Матрицы сопротивлений уравнения (1) следующие:
Суммируя первые два строки уравнения (1), где
Матрицы индуктивности изменены как
Уравнения напряжения и магнитной связи для закороченных витков ( β s 2 ) равны
Выражение электромагнитного момента может быть выражено в машинном abc переменные:
Матрицы индуктивности задаются формулами (9) — (11)
3.Анализ компонентов последовательности и извлечение параметров
Симметричные компоненты — надежный инструмент для анализа и решения проблем любой несбалансированной системы. Симметричные компоненты являются надежными индикаторами повреждений статора. В принципе, симметричные (исправные) двигатели, питаемые от симметричных трехфазных источников напряжения, не создают токов обратной последовательности. При возникновении короткого замыкания симметрия будет нарушать и генерировать токи обратной и нулевой последовательности. Что касается практики симметричных компонентов, три набора симметричных сбалансированных фаз выводятся из любого набора несимметричных параметров.Они распознаются как компоненты положительной, отрицательной и нулевой последовательности. Используя преобразование Фортескью, задаваемое уравнением (12), симметричные компоненты ( I P , I N , I 0 ) вычисляются из несбалансированных фазных токов ( I a , I b , I c ).
По сути, трехфазный асинхронный двигатель представляет собой симметричную систему в нормальных условиях и вырабатывает только токи прямой последовательности.Он генерирует положительную, отрицательную и нулевую последовательность, когда симметрия нарушается во время аварийной ситуации.
Программа MATLAB предназначена для создания имитационной модели трехфазного двигателя с повреждением одной из фазных обмоток. Из-за сложности создания неисправности и экспериментального измерения фазных токов для высоких значений процентного короткого замыкания мы вынуждены создать модель Simulink. Simulink асинхронного двигателя с закорачиванием обмотки статора построен на основе фундаментальных уравнений, упомянутых в разделе 2.Модель Simulink двигателя с межвитковым замыканием показана на рисунке 1.
Модель смоделирована для различных уровней короткого замыкания в однофазной обмотке, а значения фазных токов сохранены в рабочем пространстве MATLAB. По этим значениям рассчитываются ток обратной последовательности, ток прямой последовательности и токи нулевой последовательности. Фазные токи и токи последовательности для различных уровней повреждения приведены в таблице 1.
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||