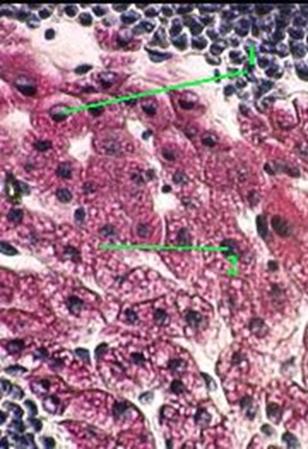
Препарат № 14 – Пигментные включения в хроматофорах кожи головастика
Препарат представляет собой неокрашенный срез (Микрофото 14). (рис. 35)
При малом увеличении видно, что структурные элементы кожи представляют как бы фон, на котором выделяются крупные пигментные клетки с отростками – меланофоры.
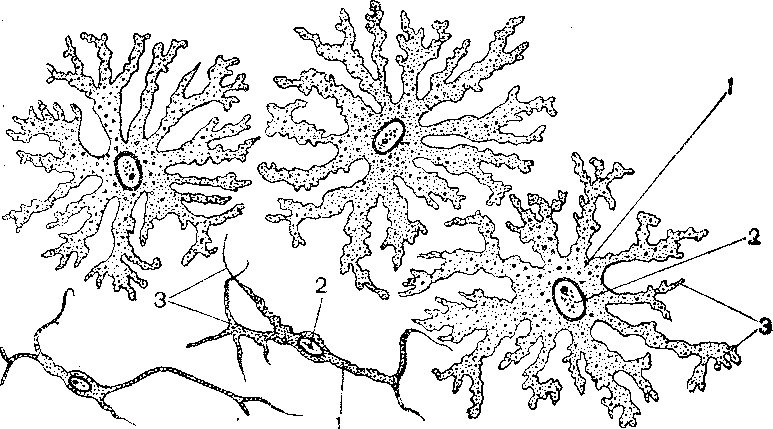 Рис. 35 Пигментные клетки-хроматофоры.
Кожа головастика. 1 – глыбки меланина,
2 – ядра, 3 – отростки меланофоров.
Рис. 35 Пигментные клетки-хроматофоры.
Кожа головастика. 1 – глыбки меланина,
2 – ядра, 3 – отростки меланофоров.
При большом увеличении в цитоплазме тел и отростков меланофоров видно значительное количество глыбок меланина, которые могут маскировать ядра этих клеток. Отростки меланофоров способны изменять длину, вследствие чего меняется количество пигментных включений на единицу объема цитоплазмы и интенсивность окраски этих клеток, а следовательно, и цвет кожи животного. Что является приспособительной, защитной реакцией организма головастика. Меланофоры встречаются в эпидермисе и соединительной ткани кожи, в сосудистой оболочке и сетчатке глаза, а также в некоторых внутренних органах человека и животных.
Обозначения: 1 – меланин, 2 – замаскированные ядра, 3 – отростки меланофоров.
Препарат № 15 –Желточные включения в бластомерах
Препарат представляет собой гистологический срез, окрашенный пиксофуксином (Микрофото 15).(рис. 36)
При малом увеличении виден зародыш лягушки на ранней стадии развития – дробления оплодотворенной яйцеклетки. Клетки, образовавшиеся в результате этого процесса, характеризуются значительной величиной и называются
 Рис. 36. Желточные включения. Бластомеры
дробящегося яйца лягушки.
Рис. 36. Желточные включения. Бластомеры
дробящегося яйца лягушки.
При большом увеличении в видно, что вся цитоплазма бластоиера заполнена желточными включениями – гранулами желтого цвета, палочковидной, округлой или овальной формы. Желточные включения образуются в яйцеклетках постепенно, в процессе их развития при непосредственном участии сетчатого аппарата Гольджи и служат питательным материалом для развивающегося зародыша. Помимо желточных гранул, в цитоплазме бластомеров встречаются мелкие буровато- коричневые пигментные включения, которые представляют собой защитное приспособление от сильного воздействия световых лучей. Скопление белка, различимые в световом микроскопе, в норме встречаются только в яйцеклетках и клетках зародыша на ранних этапах эмбриогенеза.
Обозначения: 1 – желточные включения, 2 – пигментные включения.
Контрольные вопросы
Какие отличия существуют между понятиями: протоплазма, протопласт, цитоплазма?
Какие структуры входят в состав цитоплазмы?
Что называется цитоплазматическим матриксом?
Какие функции выполняет цитоплазматический матрикс?
Какие структуры относятся к цитоскелету?
Какие белки входят в состав микрофиламентов?
Какие белки входят в состав микротрубочек?
Какие белки входят в состав промежуточных филаментов?
ЗАНЯТИЕ 5
Тема 5. НЕМЕМБРАННЫЕ ОРГАНОИДЫ
Содержание. Немембранные органоиды эукариотической клетки.
Средства наглядности. Таблицы.
Оборудование и материалы.
Задания для аудиторной работы
1. Законспектируйте теоретическую часть занятия. Обратите внимание на термины, выделенные курсивом.
2. Ответьте на контрольные вопросы.
3. Заполните таблицу «Немембранные органоиды эукариотической клетки»
Задания для внеаудиторной работы
Продолжите изучение основных классов органических соединений клетки. Обратите внимание на структуру и функции нуклеиновых кислот и белков.
Теоретическая часть
К немембранным органоидам эукариотической клетки относятся органоиды, не имеющие собственной замкнутой мембраны, а именно: рибосомы и органоиды, построенные на основе тубулиновых микротрубочек – клеточный центр и органоиды движения (жгутики и реснички).
Рибосомы
Рибосомы – немембранные органоиды, обеспечивающие биосинтез белков (полипептидов) с генетически обусловленной структурой.
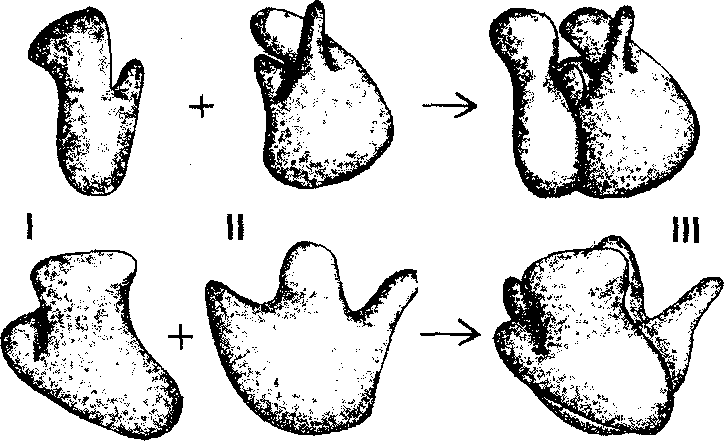 Рис.
37. Рибосома I
— малая субъединица, Il — большая субъединица, III — объединение субъединиц; верхний и
нижний ряды — изображения в разных проекциях, {По Б.
Албертсу и соавт., с изменениями).
Рис.
37. Рибосома I
— малая субъединица, Il — большая субъединица, III — объединение субъединиц; верхний и
нижний ряды — изображения в разных проекциях, {По Б.
Албертсу и соавт., с изменениями).
Рибосомы в комплексе с внутриклеточными мембранами впервые выделил Альберт Клод (1940). В 1956-1958 гг. рибосомы были выделены в чистом виде, а Р.Б. Робертс (1958) предложил сам термин «рибосома». В 1955-1959 гг. было доказано, что на рибосомах синтезируются полипептиды. Изучение структуры рибосом практически завершилось в 1980-е гг.
Рибосомы – компактные частицы диаметром 25-30 нм, состоящие из двух субъединиц с соотношением масс примерно 2:1. Для объединения субъединиц в целостную структуру необходимы ионы магния. В клетке содержится несколько десятков тысяч рибосом, при этом их количество сильно варьирует в зависимости от физиологической активности клетки.
Большинство рибосом в клетке находится в цитоплазматическом матриксе, причем существует равновесие: рибосома малая субъединица + большая субъединица.
При биосинтезе белка малая субъединица рибосомы с помощью специфических белков (факторов инициации) и метиониновой тРНК образует инициирующий комплекс, узнает начало мРНК, присоединяется к ней и скользит до точки начала инициации биосинтеза белка (как правило, это кодон АУГ).
Затем происходит присоединение большой субъединицы. При объединении субъединиц образуется целостная рибосома, которая несет два активных центра: А–участок (аминоацильный, служит для присоединения тРНК с соответствующей аминокислотой) и Р–участок (пептидилтрансферазный, служит для образования пептидной связи между аминокислотами).
После терминации (окончания) биосинтеза белка рибосома под воздействием белковых факторов вновь разделяется на субъединицы.
При биосинтезе белка обычно образуются полисомы – комплексы из одной молекулы мРНК и множества рибосом. Полисомы могут располагаться в матриксе цитоплазмы или прикрепляться к поверхности гранулярной эндоплазматической сети. Полисомы цитоплазматического матрикса осуществляют синтез белков, остающихся в клетке. Полисомы гранулярной эндоплазматической сети синтезируют как клеточные белки, так и экспортные белки. Экспортные белки поступают в полость эндоплазматической сети через поры (участки мембран, содержащие белки–переносчики), модифицируются и, пройдя через аппарат Гольджи, выводятся из клетки в составе секреторных пузырьков путем экзоцитоза.
Известно два основных типа рибосом: прокариотический тип и эукариотический тип.
Таблица 12.
ПРЕПАРАТ № 14 – Пигментные включения в хроматофорах кожи головастика
⇐ ПредыдущаяСтр 17 из 44Следующая ⇒Препарат представляет собой неокрашенный срез (Микрофото 14). (рис. 35)
При малом увеличении видно, что структурные элементы кожи представляют как бы фон, на котором выделяются крупные пигментные клетки с отростками – меланофоры.
Рис. 35 Пигментные клетки-хроматофоры. Кожа головастика. 1 – глыбки меланина, 2 – ядра, 3 – отростки меланофоров.
При большом увеличении в цитоплазме тел и отростков меланофоров видно значительное количество глыбок меланина, которые могут маскировать ядра этих клеток. Отростки меланофоров способны изменять длину, вследствие чего меняется количество пигментных включений на единицу объема цитоплазмы и интенсивность окраски этих клеток, а следовательно, и цвет кожи животного. Что является приспособительной, защитной реакцией организма головастика. Меланофоры встречаются в эпидермисе и соединительной ткани кожи, в сосудистой оболочке и сетчатке глаза, а также в некоторых внутренних органах человека и животных.
Обозначения: 1 – меланин, 2 – замаскированные ядра, 3 – отростки меланофоров.
ПРЕПАРАТ № 15 –Желточные включения в бластомерах
Препарат представляет собой гистологический срез, окрашенный пиксофуксином (Микрофото 15).(рис. 36)
При малом увеличении виден зародыш лягушки на ранней стадии развития – дробления оплодотворенной яйцеклетки. Клетки, образовавшиеся в результате этого процесса, характеризуются значительной величиной и называются бластомерами.
Рис. 36. Желточные включения. Бластомеры дробящегося яйца лягушки.
При большом увеличении в видно, что вся цитоплазма бластоиера заполнена желточными включениями – гранулами желтого цвета, палочковидной, округлой или овальной формы. Желточные включения образуются в яйцеклетках постепенно, в процессе их развития при непосредственном участии сетчатого аппарата Гольджи и служат питательным материалом для развивающегося зародыша. Помимо желточных гранул, в цитоплазме бластомеров встречаются мелкие буровато- коричневые пигментные включения, которые представляют собой защитное приспособление от сильного воздействия световых лучей. Скопление белка, различимые в световом микроскопе, в норме встречаются только в яйцеклетках и клетках зародыша на ранних этапах эмбриогенеза.
Обозначения: 1 – желточные включения, 2 – пигментные включения.
Контрольные вопросы
1. Какие отличия существуют между понятиями: протоплазма, протопласт, цитоплазма?
2. Какие структуры входят в состав цитоплазмы?
3. Что называется цитоплазматическим матриксом?
4. Что есть общего между золем и гелем?
5. Какие функции выполняет цитоплазматический матрикс?
6. Какие структуры относятся к цитоскелету?
7. Какие белки входят в состав микрофиламентов?
8. Какие белки входят в состав микротрубочек?
9. Какие белки входят в состав промежуточных филаментов?
ЗАНЯТИЕ 5
Тема 5. НЕМЕМБРАННЫЕ ОРГАНОИДЫ
Содержание. Немембранные органоиды эукариотической клетки.
Средства наглядности. Таблицы.
Оборудование и материалы. Гистологические материалы. Микроскопы.
Задания для аудиторной работы
1. Законспектируйте теоретическую часть занятия. Обратите внимание на термины, выделенные курсивом.
2. Ответьте на контрольные вопросы.
3. Заполните таблицу «Немембранные органоиды эукариотической клетки»
Задания для внеаудиторной работы
Продолжите изучение основных классов органических соединений клетки. Обратите внимание на структуру и функции нуклеиновых кислот и белков.
Теоретическая часть
К немембранным органоидам эукариотической клетки относятся органоиды, не имеющие собственной замкнутой мембраны, а именно: рибосомы и органоиды, построенные на основе тубулиновых микротрубочек – клеточный центр и органоиды движения (жгутики и реснички).
Рибосомы
Рибосомы – немембранные органоиды, обеспечивающие биосинтез белков (полипептидов) с генетически обусловленной структурой.
Рис. 37. РибосомаI — малая субъединица, Il — большая субъединица, III — объединение субъединиц; верхний и нижний ряды — изображения в разных проекциях, {По Б. Албертсу и соавт., с изменениями).
Рибосомы в комплексе с внутриклеточными мембранами впервые выделил Альберт Клод (1940). В 1956-1958 гг. рибосомы были выделены в чистом виде, а Р.Б. Робертс (1958) предложил сам термин «рибосома». В 1955-1959 гг. было доказано, что на рибосомах синтезируются полипептиды. Изучение структуры рибосом практически завершилось в 1980-е гг.
Рибосомы – компактные частицы диаметром 25-30 нм, состоящие из двух субъединиц с соотношением масс примерно 2:1. Для объединения субъединиц в целостную структуру необходимы ионы магния. В клетке содержится несколько десятков тысяч рибосом, при этом их количество сильно варьирует в зависимости от физиологической активности клетки.
Большинство рибосом в клетке находится в цитоплазматическом матриксе, причем существует равновесие: рибосома малая субъединица + большая субъединица.
При биосинтезе белка малая субъединица рибосомы с помощью специфических белков (факторов инициации) и метиониновой тРНК образует инициирующий комплекс, узнает начало мРНК, присоединяется к ней и скользит до точки начала инициации биосинтеза белка (как правило, это кодон АУГ).
Затем происходит присоединение большой субъединицы. При объединении субъединиц образуется целостная рибосома, которая несет два активных центра: А–участок (аминоацильный, служит для присоединения тРНК с соответствующей аминокислотой) и Р–участок (пептидилтрансферазный, служит для образования пептидной связи между аминокислотами).
После терминации (окончания) биосинтеза белка рибосома под воздействием белковых факторов вновь разделяется на субъединицы.
При биосинтезе белка обычно образуются полисомы – комплексы из одной молекулы мРНК и множества рибосом. Полисомы могут располагаться в матриксе цитоплазмы или прикрепляться к поверхности гранулярной эндоплазматической сети. Полисомы цитоплазматического матрикса осуществляют синтез белков, остающихся в клетке. Полисомы гранулярной эндоплазматической сети синтезируют как клеточные белки, так и экспортные белки. Экспортные белки поступают в полость эндоплазматической сети через поры (участки мембран, содержащие белки–переносчики), модифицируются и, пройдя через аппарат Гольджи, выводятся из клетки в составе секреторных пузырьков путем экзоцитоза.
Известно два основных типа рибосом: прокариотический тип и эукариотический тип.
Таблица 12.
Типы рибосом
| Критерии Для сравнения | Типы рибосом | |
| Прокариотический тип | Эукариотический тип | |
| Нахождение в клетках | Рибосомы эубактерий и цианобактерий Пластидные рибосомы Митохондриальные Рибосомы Рибосомы архебактерий (сходны с эукариотическими) | Цитоплазматические Рибосомы эукариот: Животных, Грибов И растений |
| Соотношение РНК : белок | 2 : 1 | 1 : 1 |
| Количество рРНК в малой субъединице | 1 молекула длиной 1500-1600 нуклеотидов (минимум – 950 в митохондриальных рибосомах) | 1 молекула длиной около 1800 нуклеотидов |
| Количество рРНК в большой субъединице | 2 молекулы: 1 молекула длиной около 3000 нуклеотидов, 1 молекула длиной около 120 нуклеотидов | 3 молекулы: 1 молекула длиной около 4-5 тысяч нуклеотидов, 1 молекула длиной около 120 нуклеотидов, 1 молекула длиной около 160 нуклеотидов (эта молекула сходна с одним из участков прокариотической рРНК) |
| Содержание белка (почти все молекулы уникальны) | Примерно 20 молекул в малой субъединице и 30 молекул – в большой | Примерно 30 молекул в Малой субъединице и 40 молекул – в большой |
Клеточный центр
Клеточный центр – органоид, контролирующий образование и рост микротрубочек цитоскелета эукариотической клетки. (рис. 38)
Рис. 38.Клеточный центр1 – триплеты микротрубочек; 2 – радиальные спицы; 3 – центральная структура «колеса телеги»; 4 – сателлит; 5 – лизосома; 6 – диктиосомы комплекса Гольджи; 7 – окаймленный пузырек; 8 – цистерна гранулярной эндоплазматической сети; 10 – митохондрия; 11 – остаточное тельце; 12 – микротрубочки; 13 – кариотека (по Р. Крстичу, с изменениями).
Основные функции клеточного центра – формирование систем микротрубочек цитоскелета, органоидов движения, веретена деления.
Основу клеточного центра составляет пара центриолей (иногда их число достигает 4-6-8 в полиплоидных клетках), расположенных в центре клетки.
Впервые центриоли обнаружил и описал В. Флемминг (1875), но подробно они были изучены лишь с помощью электронного микроскопа. Центриоли обнаруживают тесную связь с ядерной оболочкой, а у многих низших эукариот (простейшие, водоросли, грибы, мезокариоты) центриоли встроены в оболочку ядра.
Клеточный центр всегда обнаруживается в клетках многоклеточных животных. У прокариот клеточный центр и тубулин отсутствуют. В клетках грибов, водорослей и одноклеточных животных клеточный центр обнаруживается далеко не всегда, а в клетках высших растений не обнаружен (за редким исключением). При отсутствии клеточного центра его функции у эукариот выполняет центр образования микротрубочек – ЦОМТ.
Центриоль представляет собой полый цилиндр диаметром около 0,15 мкм и длиной 0,3-0,5 мкм (реже – несколько мкм). Стенки центриолей состоят из 9 триплетов тубулиновых микротрубочек, связанных динеиновыми мостиками. Обычно центриоли располагаются парами: одна центриоль – материнская, а другая – дочерняя; дочерняя центриоль расположена перпендикулярно материнской. Такая пара центриолей – диплосома – имеет Т–образную или Г–образную форму. Диплосома окружена плотным слоем цитоплазмы, образующим центросферу, или центросому. Во время деления клетки от центросферы отходят плотные тяжи микротрубочек, образующие астросферу.
Удвоение центриолей происходит примерно в середине интерфазы: материнская и дочерняя центриоли расходятся, и от каждой центриоли отпочковывается новая центриоль. У разных групп организмов этот процесс имеет свои особенности.
Органоиды движения
К органоидам движения относятся жгутики и реснички. (рис. 39, 40) В основе органоидов движения лежит система тубулиновых микротрубочек.
У всех эукариот органоиды движения имеют сходную структуру.
Типичный жгутик состоит из базального тела (или кинетосомы), переходной зоны, главного стержня и кончика. Если жгутиков два и более, то их базальные тела соединены жгутиковыми корнями. Мелкие многочисленные жгутики (их может быть до 10-15 тысяч на клетку) называются ресничками, их базальные тела образуют единую систему (ризопласт), связанную с ядерной оболочкой, аппаратом Гольджи и митохондриями. Главный стержень и кончик жгутика или реснички покрыты мембраной, являющейся продолжением плазмалеммы. Базальные тела окружены складкой плазмалеммы.
Базальное тело представляет собой полый цилиндр, стенки которого образованы девятью триплетами тубулиновых микротрубочек, связанных динеиновыми мостиками. Таким образом, центриоль и базальное тело являются альтернативными формами одного органоида.
Рис. 39. Ресничка— тонкий вырост на поверхности клетки. Стержень реснички образован аксонемой — системой микротрубочек 9+2. В основании реснички расположено базальное тельце, служащее матрицей для формирования аксонемы {из Kopf-Maier P, Merker H~J, 1989)
А БРис 40. Характер движения жгутика (А) и реснички (Б). Несмотря на то, что молекулярная основа подвижности у жгутиков и ресничек одинакова, характер их движения различен. Жгутики обычно длиннее ресничек, и для них характерно синусоидальное движение, в отличие от циклических волнообразных изгибов ресничек [ по Satir P (1974) из Fawcett DW, (1981)].
Переходная зона находится в области пересечения жгутика с плазмалеммой. В центре переходной зоны лежит аксиальная гранула, от которой отходит две одиночные микротрубочки, которые идут вдоль оси жгутика до самого конца. На периферии переходной зоны лежит базальный диск, в котором утрачивается одна из трех микротрубочек каждого триплета, и триплеты превращаются в дублеты.
Главный стержень представлен цилиндром, стенки которого образованы девятью дублетами микротрубочек, связанных динеиновыми мостиками. Вдоль оси главного стержня тянутся две одиночные микротрубочки. Полость цилиндра заполнена собственным матриксом.
По мере приближения к кончику дублеты постепенно утрачивают одну из двух микротрубочек, а затем исчезают полностью. Заканчивается жгутик двумя центральными микротрубочками, покрытыми мембраной.
У ряда организмов (некоторые водоросли) обнаружены незначительные отклонения от типичной организации жгутиков: центральные трубочки или отсутствуют, или имеется лишь одна. В специализированных клетках (например, в сперматозоидах человека) в состав дублета входит одна полная микротрубочка (из 13 микрофиламентов) и одна неполная (из 11 микрофиламентов). У некоторых групп эукариот жгутики и реснички отсутствуют (покрытосеменные растения, нематоды, членистоногие, часть одноклеточных животных, водорослей и голосеменных растений). (рис. 41)
Рис. 41. Актиновый микрофиламент 1 – глобулы актина; 2 – тропомиозин; 3 – тропонины (по Б. Альбертсу и соавт., с изменениями).
Жгутики прокариот имеют совершенно иную организацию. В их основе лежит полый цилиндр из белка флагеллина. В состав основания прокариотического жгутика входит около 12 разных белков.
Реснички и жгутики
Ресничка представляет собой вырост клетки, окруженный плазмалеммой. У основания реснички на уровне кортикального слоя цитоплазмы находится базальное тельце (кинетосома), которое образовано девятью периферическими триплетами коротких микротрубочек, окружающих один белковый центральный цилиндр. Каждый периферический триплет соединен с ним посредством белковых «спиц».
От каждой наружной микротрубочки отходит отросток, напоминающий флажок. Над базальным тельцем в названный цитоцлазматический вырост направляется осевая нить (осевой филамент, аксонема), тоже образованная микротрубочками (строение последних описано ранее). От каждого базального тельца к соседнему (к основанию другой реснички) отходит короткий боковой отросток, а в глубь цитоплазмы — поперечно исчерченный базальный корешок (периодичность исчерченности равна 65 нм).
Рис. 42. Ресничка I — общая структура, II — срез через внеклеточную часть, III — срез через внутриклеточную часть, IV — срез через базальное тельце, 1 — корешок, 2 — базальное тельце, 3 — плазмалемма поверхности клетки, 4 — микроворсинки, окружающие ресничку, 5 — центральна? капсула, 6 — дуплеты периферических микротрубочек, 7 — центральная пара микротрубочек, 8 — плазмалемма на поверхности реснички, 9 — динеиновые «ручки», 10-радиальная спица, 11 — нексии, 12 — субьединицаА, 13- субъединица В. 14- субъединица С, 15 — центральный цилиндр (по Р. Крстичуи по Ю. С. Ченцову, с изменениями).
Сразу над базальным тельцем микротрубочки аксонемы тоже образуют девять периферических триплетов, но над уровнем плазмалеммы в периферических комплексах одна из микротрубочек редуцируется; в центральной группе появляется пара микротрубочек, окруженная белковой оболочкой, имеющей вид цилиндра. Поэтому на большей части протяжения реснички тянутся дуплеты микротрубочек. Девять дуплетов располагаются на периферии, один — в центре.
В результате на поперечном сечении реснички ее возвышающаяся часть напоминает колесо с девятью спицами, в центре которого лежит нечетко контурирующаяся центральная капсула, окружающая две центральные одиночные микротрубочки, с которыми она связана белковыми отростками. Периферические дуплеты состоят из двух микротрубочек (А и В) и окружают центральную капсулу (см. рис. 41).
От микротрубочки А к центральной капсуле отходит радиальная «спица» с периодичностью расположения вдоль аксонемы, равной 29 нм. По направлению к соседней трубочке В отходит пара «ручек», образованных, как и «спицы», белком динеином с периодичностью 24 нм. Соседние дуплеты микротрубочки соединены между собой с периодичностью 86 нм поперечными мостиками, образованными белком нексином.
Реснички являются производными не только поверхностного комплекса клетки, но и клеточного центра. В начале их развития происходит многократная редупликация цен-триолей. Новые центриоли парами мигрируют к поверхности клетки. Здесь происходит их модификация.
Одна из центриолей (проксимальная) ложится в поверхностном комплексе вблизи от плазмалеммы у основания будущей реснички. Другая центриоль (дистальна] ложится между ней и плазмалеммой. Триплеты микротрубочек дистальной центриоли становятся дуплетами, в проксимальной центриоли триплеты сохраняются. В клеточном центре идет сборка микротрубочек, они направляются к дистальной центриоли и наращивают ее длину. Дистальная центриоль растет и влечет за собой плазмалемму. Рост аксонемы в длину происходит за счет полимеризации тубулина у дистальных концов двух внутренних трубочек каждого триплета базального тельца.
Жгутик (flagellum) эукариотической клетки (например, сдермия) напоминает ресничку, но он длиннее.
Реснички и жгутики выполняют функцию движения. Все реснички клетки совершают координированные колебания. Это достигается посредством скольжения дуплетов микротрубочек относительно друг друга. Оно обусловлено изменениями конфигурации молекул белка динеина- Динеин обладает аденозинтрифосфатазной активностью. При гидролизе АТФ выделяется свободная энергия, за счет которой динеиновые ручки выпрямляются, контактируют с соседним дуплетом микротрубочек и сдвигают его по направлению к верхушке реснички. При регенерации АТФ ручки отделяются от соседнего дуплета и опускаются вниз к основанию реснички.
Все реснички клетки совершают координированные колебательные движения. Они похожи на движения рук пловца брассом. Сначала ресничка резко наклоняется над поверхностью клетки. При этом слизь, которой обычно здесь покрыта поверхность, прогоняется в направлении наклона. Далее наклоненная ресничка совершает поворот на 180°. Поскольку она скользит параллельно поверхности клетки, слизь над ней назад не возвращается. Затем ресничка снова выпрямляется и перемещает слизь. Потом начинается новый цикл.
Число ресничек достигает нескольких сотен. Так, до 250 ресничек длиной 5—15 мкм и диаметром 0,15—0,25 мкм покрывают апикальную поверхность реснитчатых эпителиоцитов верхних дыхательных путей, маточных труб, семявыводящих канальцев.
ПРАКТИЧЕСКАЯ ЧАСТЬ
2.9. Цитоплазматические включения
Включения – это непостоянные компоненты клетки, представляющие собой отложения веществ, не участвующие в данный момент в клеточном метаболизме. Большинство включений видимы в световой микроскоп в виде гранул и располагаются в гиалоплазме, органоидах, либо в вакуолях. Существуют твердые и жидкие включения. В зависимости от нужд клетки, они могут присутствовать в больших количествах или исчезать.
Любые твёрдые включения в клетке, как правило, максимально обезвожены. Этот феномен не только снижает объём включений, но и препятствует развитию в них бактерий. Кроме того, в процессе образования твёрдые включения становятся химически инертными, что не позволяет им вступать в химические реакции с клеточными соединениями.
По химическому составу различают углеводные, белковые, липидные и минеральные включения. По функции — трофические, секреторные, специальные и т.д.
а. Трофические (питательные) включения
К ним относятся белковые, жировые и полисахаридные включения.
Белковые включения. В клетке есть соединения, важность которых определяется тем, что в случае нужды они могут стать предшественниками целого ряда других, жизненно необходимых, для клетки веществ. К таким соединениям относятся аминокислоты. Они могут использоваться в клетке в качестве источников энергии для синтеза углеводов, жиров, гормонов и других метаболитов. Поэтому белковые включения фактически представляют собой своеобразное клеточное сырьё для производства аминокислот.
Судьба белковых включений во всех клетках примерно одинакова. Прежде всего они сливаются с лизосомой, где специальные ферменты расщепляют белки до аминокислот. Последние выходят из лизосом в цитоплазму. Часть из них в цитоплазме взаимодействует с тРНК и в таком виде транспортируется к рибосомам на синтез белка. Другая часть вступает в специальные биохимические циклы, где из них синтезируются жиры, углеводы, гормоны и другие метаболиты. И наконец, аминокислоты принимают участие в энергетическом обмене клетки.
Полисахаридные включения. Для животных клеток и клеток грибов — основным запасным питательным включением является гликоген. Для растений таким включением является крахмал.
Гликоген у человека в основном депонируется в клетках печени и используется не только для нужд самой клетки, но и в качестве энергетических ресурсов для всего организма. В последнем случае гликоген расщепляется в клетке до глюкозы, которая выходит из клетки в кровь и разносится по организму.
Гликоген – это большая ветвистая молекула, состоящая из глюкозных остатков. Специальные внутриклеточные процессы при необходимости отщепляют от молекулы гликогена глюкозные остатки и синтезируют глюкозу. Последняя поступает в кровь и расходуется на нужды клетки. Казалось бы, проще запасти в клетке саму глюкозу, не превращая её в гликоген, тем более что молекула глюкозы растворима и достаточно быстро проходит в клетку через плазматическую мембрану. Однако этому мешает то, что глюкоза также быстро, не задерживаясь, выходит из клетки. Задержать её в клетке в чистом виде практически невозможно. Кроме того, депонирование глюкозы в больших количествах опасно, т.к. это может привести к созданию такого градиента концентраций, что вначале произойдёт набухание клетки из-за поступления воды, а затем и её гибель. Поэтому специальная система ферментов, слегка видоизменив молекулу глюкозы, связывает её с такой же молекулой. Создаётся гигантская ветвистая молекула, состоящая из глюкозных остатков – гликоген. Эта молекула уже нерастворима, как глюкоза, и не способна изменить осмотические свойства клетки.
Жировые включения.Эти включения в гиалоплазме могут находиться в виде капель. Многие растения содержат масла, например подсолнечник, арахис и т.д. Богата жировыми включениями жировая ткань человека, служащая для предохранения организма от потерь тепла, энергетическим депо и как амортизатор при механических воздействиях.
Необходимо отметить, что запасов гликогена в организме среднего взрослого человека достаточно на один день нормальной активности, тогда как запаса жиров хватит на месяц. Если бы главным энергетическим резервом в нашем организме был гликоген, а не жиры, вес тела увеличился бы в среднем на 25 кг.
В некоторых случаях появление жировых включений в клетке является тревожным сигналом неблагополучия. Так, при заболевании дифтерией токсин микроорганизма блокирует утилизацию жирных кислот и они накапливаются в больших количествах в цитоплазме. При этом нарушается метаболизм и клетка гибнет. Чаще всего такие нарушения происходят в клетках сердечной мышцы. Заболевание называется дифтерийный миокардит.
Все питательные включения используются клеткой в моменты интенсивной жизнедеятельности. В эмбриогенезе существует потребность в большом количестве питательных веществ. Поэтому ещё на стадии овогенеза яйцеклетка интенсивно запасает различные питательные вещества (желток и др.) в виде включений, которые обеспечивают прохождение первых этапов эмбрионального развития.
б. Секреторные включения
Различные секреторные гранулы, образующиеся в железистых клетках животных, разнообразны по химической природе и могут быть представлены ионами, ферментами, гормонами, гликопротеинами и т.д., например, пищеварительные ферменты, синтезируемые клетками поджелудочной железы. Сигналом к формированию и опорожнению секреторных включений в поджелудочной железе является приём пищи. До приёма пищи происходит накопление включений в цитоплазме. Определяя количество включений в клетках поджелудочной железы, можно примерно предположить, чьи это клетки – голодного или сытого человека.
в. Пигментные включения
Обеспечивают окраску тканей и органов. В клетках животных часто запасается пигмент меланин. У растений — каротиноиды, флавоноиды.
г. Экскреторные включения
У растений, не имеющих специальных выделительных органов, нередко в вакуолях накапливаются конечные продукты жизнедеятельности в виде солей оксалата или карбоната кальция. В дальнейшем растение сбрасывает листья или кору вместе с такими накопленными кристаллическими включениями.










Организация и функции цитоскелета
Топография структур (где находятся) | Связь с органоидами Клетки | Основные Функции |
Микрофиламенты | ||
Микротрубочки | ||
Практическая часть
Изучение деления клетки путем митоза. Фиксация корней к занятию .
Задания
1.Изучите организацию и функцию цитоскелета.
Препарат № 2 – митоз в корешке лука.
Препарат представляет собой разрез корешка лука, окрашенный железным гематоксилином (Микрофото 2).
При малом увеличении видно, что ткань корешка лука построена из продолговатых клеток с хорошо различимыми оболочкой и ядром.
При большом увеличении находим клетки, в ядра которых не изменены. Передвигая препарат, отыскиваем клетки, где хромосомы обнаруживаются нити-хромосомы, а так же клетки, где хромосомы обнаруживаются вместо ядра. Встречаются клетки на стадии материнской звезды, в которых хромосомы располагаются по экватору ахроматинового веретена, а также клетки с фигурой дочерней звезды ( хромосомы расходятся по полюсам клетки). При большом увеличении зарисовать отдельные клетки.
Обозначения: 1ядро. 2 – клетки, 3 – ядрышки. 4 – цитоплазма. 5 – плазматическая мембрана. 6 – зона роста. 7 – целлюлозная выстилка.
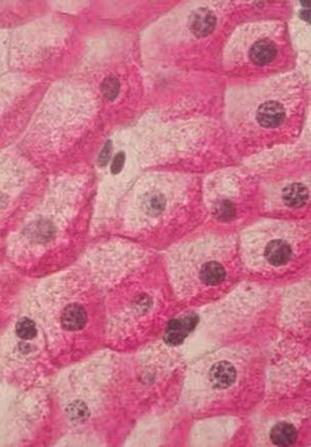
Препарат № 12 – Липидные (жировые) включения в клетках печени аксолотля.
Препарат представляет собой срез печени аксолотля, окрашенного Суданом III(Микрофото 12) .(рис. 33)
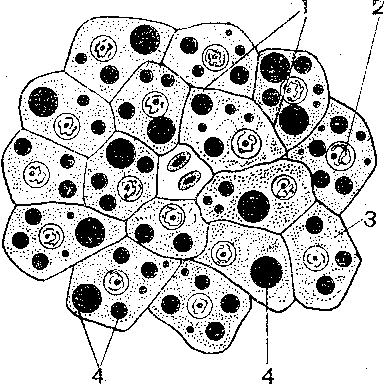
Рис. 33.Жировые включения в клетках печени аксолотля. 1 – клеточные границы гепатоцитов, 2 – круглые структурированные ядра, 3 – цитоплазма, 4 – жировые включения.
При малом увеличении производим ориентировку препарата и переводим микроскоп на большое увеличение. Ядра печеночных клеток – гепатоцитов округлые, окрашены в красный цвет. В цитоплазме видны (липидные) включения в виде шариков различной величены, которые окрашены в черный цвет. Между клетками видны границы – клеточные оболочки.
Обозначения: 1 – клеточная оболочка, 2 – ядро, 3 – цитоплазма, 4 – липидные включения.
Препарат № 13 – Включения гликогена в клетках печени аксолотля.
Препарат окрашен кармин-гематоксилином по Бесту (Микрофото 13).(рис. 34)
Гликоген является широко распространенным видом углеводных включений в животных клетках. Рассматривая препарат при малом увеличении микроскопа, видим, что печеночные клетки имеют многогранную форму. Ядра клеток округлые либо овальные, окрашены в темно – синий цвет. Цитоплазма клеток в большей или меньшей мере заполнена включениями гликогена, имеющими форму глыбок разной величены, окрашенных в красный цвет. Зарисовать при большом увеличении печеночные клетки. В цитоплазме показать включение гликогена.
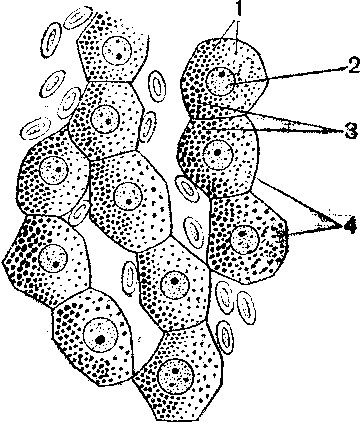 Рис. 34. Включения гликогена в клетках
печени аксолотля. 1 – гликоген в виде
глыбок, 2 – структурированные ядра, 3 –
скопление гликогена в гепатоцитах, 4 –
неокрашенные вакуоли различной величины.
Рис. 34. Включения гликогена в клетках
печени аксолотля. 1 – гликоген в виде
глыбок, 2 – структурированные ядра, 3 –
скопление гликогена в гепатоцитах, 4 –
неокрашенные вакуоли различной величины.
Обозначения: 1 –печеночные клетки, 2 – ядра, 3 – цитоплазма, 4 – включения гликогена.
Включения цитоплазмы
Включения — это непостоянные (необязательные) структурные элементы цитоплазмы.
Они заметны при световой микроскопии при общих методах окрашивания, иногда при малом и среднем увеличении, а часть из них можно выявить лишь специальными (гистохимическими, иммунологическими) методами или при электронной микроскопии. В зависимости от активности клетки, гормональных и метаболических влияний, особенностей дифференцировки, возраста, действия разнообразных факторов внешней среды в клетках можно обнаружить большое разнообразие включений по составу и количеству.
Включения указывают на особенности метаболизма, дифференцировки, функциональной активности клеток. Много включений появляется при дистрофических нарушениях в клетке, что сопровождается изменениями в ее жизнедеятельности вплоть до гибели. Иногда содержимое включений не только показатель функции, но основание для названия клетки: пигментные клетки — меланоциты; эозинофильные, базофильные и нейтрофильные гранулоциты крови и др.
При всем многообразии включений их можно объединить по их функциональному назначению.
Секреторные включения. Представляют собой секреторные гранулы, которые выделяются из клетки путем экзоцитоза. По химическому составу их подразделяют на белковые (серозные), жировые (липидные, или липосомы), слизистые (содержат мукополисахариды) и др. Количество включений зависит от функциональной активности клетки, стадии секреторного цикла, степени зрелости клетки. Особенно много гранул в дифференцированных, функционально активных клетках в фазу накопления секреторного цикла.
Секреторные включения образуются в комплексе Гольджи. Перед этим они проходят стадию синтеза в гр. или глад. ЭПС, реже это происходит в других структурах.
Секреторные белковые включения разнообразны по своим размерам, распределению в цитоплазме, электронной плотности. Они окружены клеточной мембраной. Полипептидные цепочки содержимого секреторных включений синтезируются в гр. ЭПС, а созревают в комплексе Гольджи. В связи с этим у клеток, синтезирующих секреторные белки, хорошо развиты данные органеллы, крупное ядро и ядрышки. Однако если клетка прекращает синтез включений, их накопление сопровождается инволюцией гр. ЭПС и комплекса Гольджи.
В экзокринных железах секреторные включения преобладают в апикальной части клетки, предполагая выведение секрета во внешнюю среду. Секреторные включения эндокринных желез концентрируются вблизи кровеносных сосудов или равномерно распределены в цитоплазме.
Слизистые секреторные включения находятся в основном в клетках слизистых секреторных желез. Примером одноклеточных секреторных желез служат бокаловидные клетки тонкой кишки. При световой микроскопии с помощью ШИК-реакции слизь хорошо видна в крупных вакуолях.
Секреторные включения, содержащие жиры (липосомы), имеются в цитоплазме сальных желез и эндокринных клеток, синтезирующих стероидные гормоны (производные холестерина). Стероидные гормоны — это мужские и женские половые гормоны, гормоны стресса (глюкокортикоиды) и гормон, контролирующий содержание ионов натрия в организме (альдостерон). В этих клетках хорошо развита глад, и гр. ЭПС, комплекс Гольджи, много митохондрий. Митохондрии эндокриноцитов участвуют в синтезе стероидных гормонов и имеют специфические особенности строения. Это крупные митохондрии с мультивезикулярными (трубчатыми) кристами.
Также выделяют секреторные включения, содержащие производные аминокислот и других аминов: норадреналин и адреналин, серотонин (мелатонин) и др.
Разнообразен состав секреторных включений в тучной клетке (лаброците) и базофильном гранулоците (базофиле). Эти клетки содержат многочисленные крупные секреторные включения, окрашивающиеся основными красителями и нередко изменяющие их оттенок. Такая способность изменять цвет красителя называется метахромазией. При электронной микроскопии видно, что в лаброцитах и гранулоцитах много крупных гранул округлой формы, различной электронной плотности.
Количество включений зависит от стадии секреторного цикла. Максимально их количество на стадии
Хроматин
Хроматин – комплекс ядерной ДНК с белками (гистоны, негистоновые белки).
Гетерохроматин (транкрипционно неактивный, конденсированный хроматин) интерфазного ядра. В СМ (видны в световой микроскоп)– базофильные глыбки, в ЭМ (видно в электронный микроскоп) – скопления плотных гранул. Располагается преимущественно по периферии ядра и вокруг ядрышек. Типичный пример гетерохроматина —тельце Барра.
Тельце Барра
Во всех соматических клетках генетически женского организма одна из Х –хромосом инактивирована и известна как половой хроматин (тельце Барра). Инактивация Х-хромосомы известна каклайонизация.
Лайонизация – механизм компенсации дозы генов Х-хромосомы у женщин объясняет гипотизаМэри Лайон.
Согласно гипотезе, инактивация Х-хромосомы происходит в раннем эмриогенезе, осуществляеися случайным образом (инактивированной может быть либо отцовская, либо материнская Х-хромосома), затрагивает целиком всю Х-хромосому и характеризуется устойчивостью, передаваясь клеточным потомкам. Клетки женского организма по экспрессии генов Х-хромосомы мозаичны.
Эухроматин – транскрипционно активная и менее конденсированная часть хроматина, локализуется в более светлых участках ядра между гетерохроматином.
Хромосома
Хромосомы видны при митозе или мейозе, когда хроматин конденсирован полностью.
ДНК
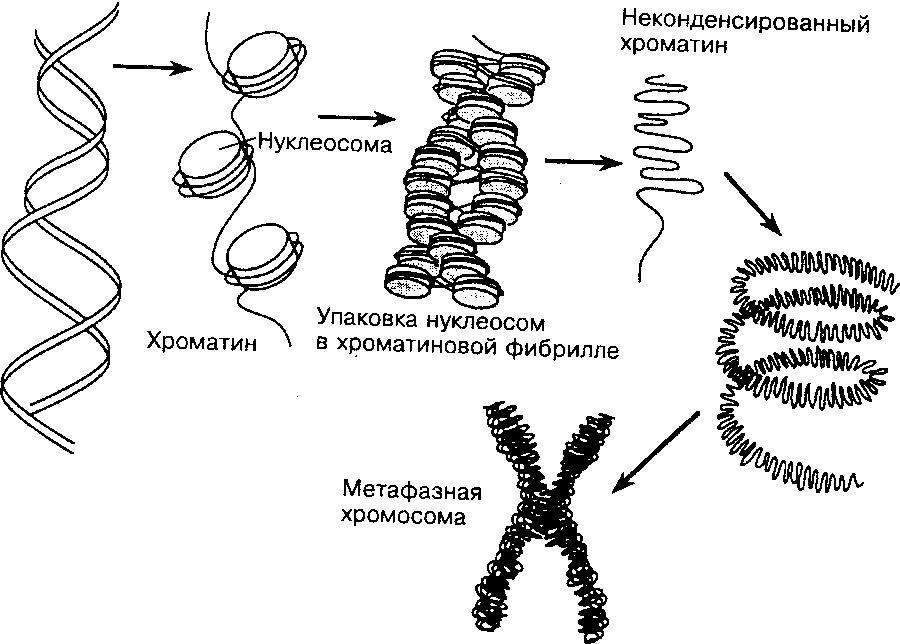
Рис. 8. Организация хроматина в хромосоме. Хроматин состоит из структурных единиц — нуклеосом, разделённых интервалами в 200 пар оснований. Во время митоза в результате плотной упаковки нуклеосом хроматин полностью конденсируется, формируя видимые хромосомы (из Widneil СС, Pfeninger КН, 1990)
ОРГАНИЗАЦИЯ ХРОМАТИНА В ХРОМОСОМЕ
Хроматин состоит из структурных единиц – нуклеосом, разделенных интервалами в 200 пар оснований. Во время митоза в результате плотной упаковки нуклеосом хроматин полностью конденсируется, формируя видимые хромосомы. (рис. 8)
Состав хромосом
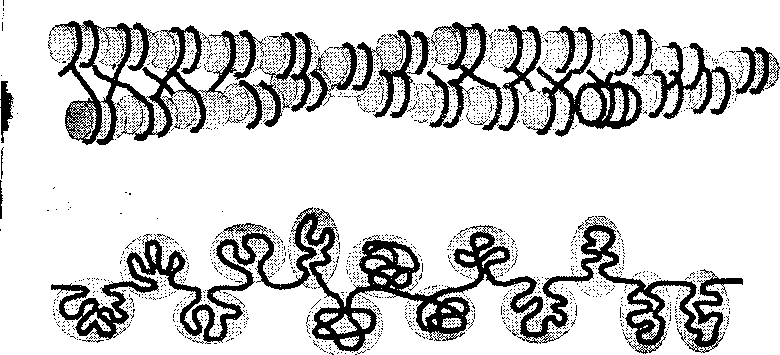
Каждая хромосома содержит одну молекулу ДНК и ДНК- связывающие белки; хроматин в составе хромосомы образует многочисленные петли. Хромосома состоит из структурных единиц – нуклеосом. (рис. 9, 10)
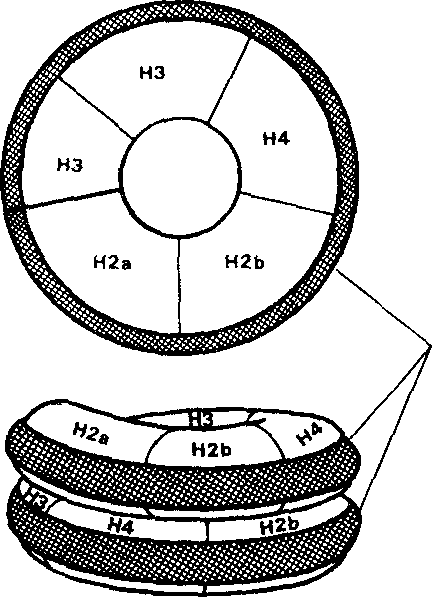
Рис.9. Нуклеосома в неконденсированном хроматине содержит по две копии гистонов Н2А, Н2В, НЗ и Н4. Двойная спираль ДНК лежит на поверхности октамера гистонов и накручена на него. В конденсированном хроматине дополнительно присутствует гистон H1, соединяющий нуклеосомы [из Trifonov EN, 1981|
Нуклеосомы – сферические структуры диаметром 10 нм.
 I
I
II
 III
III
 IV
IV
 V
V
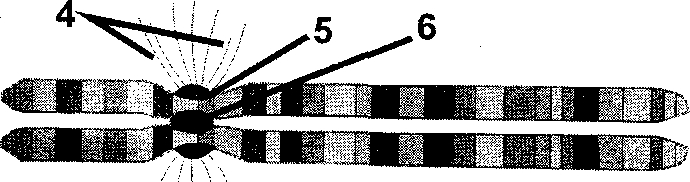
Рис. 10. Уровни упаковки ДНК в хромосоме I– нуклеиновая нить,II– хроматиновая фибрилла,III– серия петельных доменов,IV– конденсированный хроматин в составе петельного домена,V– метафазная хромосома; 1 – гистон Н1, 2 – ДНК, 3 – прочие гистоны, 4 – микротрубочки ахроматинового веретена, 5 – кинетохор, 6 – центромера, 7 – хроматиды (по Б. Албертсу и соавт., с изменениями и дополнениями).
ГЕНОМ
Геном– полный комплект генов в хромосомах.Кариотип – описывает количество и структуру хромосом.Гаплоидный набор –23 хромосомы – характерен для гамет.Диплоидный набор — стандарт хромосом (23 х 2) – для соматических клеток.
Митоз в растительных клетках корешка лука. Окраска железным гематоксилином
Обратная связь
ПОЗНАВАТЕЛЬНОЕ
Сила воли ведет к действию, а позитивные действия формируют позитивное отношение
Как определить диапазон голоса — ваш вокал
Как цель узнает о ваших желаниях прежде, чем вы начнете действовать. Как компании прогнозируют привычки и манипулируют ими
Целительная привычка
Как самому избавиться от обидчивости
Противоречивые взгляды на качества, присущие мужчинам
Тренинг уверенности в себе
Вкуснейший «Салат из свеклы с чесноком»
Натюрморт и его изобразительные возможности
Применение, как принимать мумие? Мумие для волос, лица, при переломах, при кровотечении и т.д.
Как научиться брать на себя ответственность
Зачем нужны границы в отношениях с детьми?
Световозвращающие элементы на детской одежде
Как победить свой возраст? Восемь уникальных способов, которые помогут достичь долголетия
Как слышать голос Бога
Классификация ожирения по ИМТ (ВОЗ)
Глава 3. Завет мужчины с женщиной

Оси и плоскости тела человека — Тело человека состоит из определенных топографических частей и участков, в которых расположены органы, мышцы, сосуды, нервы и т.д.
 Отёска стен и прирубка косяков — Когда на доме не достаёт окон и дверей, красивое высокое крыльцо ещё только в воображении, приходится подниматься с улицы в дом по трапу.
Отёска стен и прирубка косяков — Когда на доме не достаёт окон и дверей, красивое высокое крыльцо ещё только в воображении, приходится подниматься с улицы в дом по трапу.
 Дифференциальные уравнения второго порядка (модель рынка с прогнозируемыми ценами) — В простых моделях рынка спрос и предложение обычно полагают зависящими только от текущей цены на товар.
Дифференциальные уравнения второго порядка (модель рынка с прогнозируемыми ценами) — В простых моделях рынка спрос и предложение обычно полагают зависящими только от текущей цены на товар.
На препарате на малом увеличении видны клетки кончика корешка лука
Они
-прямоугольной формы и
— плотно расположены
При большом увеличении можно увидеть, что большинство клеток, находятся на различных стадиях деления и в интерфазе.
В интерфазных клетках хорошо выявляются структуры ядра —
-глыбки хроматина ,
-ядрышки ,
-ядерная оболочка Часть клеток находится в состоянии митоза.
Когда клетка проходит стадию профазы:
— в ядре выявляется конденсированный хроматин
— ядерная оболочка ещё сохраняется и ограничивает клубок хроматина
— на более поздних этапах оболочка распадается на фрагменты, и нити хроматина располагаются по всей клетке
Клетка — на стадии метафазы:
— ядерная оболочка исчезла,
— хромосомы образуют экваториальную пластинку
На другом участке того же препарата мы встречаем клетки на последующих стадиях митоза.
клетка — на стадии анафазы:
— хромосомный наборы расходятся по полюсам клетки.
клетка — на стадии телофазы:
-хромосомы деспирализовались, образова- лись ядерные оболочки и
— произошла цитотомия.
Центросомы в яйцеклетке лошадиной аскариды.
Окраска: железный гематоксилин.
Препарат представляет собой срез матки лошадиной аскариды. Видны яйцеклетки, лежащие в полости матки. Они находятся на разных стадиях развития.
Яйцеклетки окружены оболочкой, внутри которой происходит дробление. Найти яйцеклетку, находящуюся в метафазе, либо в анафазе первого дробления. В это время отсутствует ядерная оболочка, и хромосомы либо располагаются по экватору клетки, либо уже начали движение к полюсам. На этих стадиях на полюсах яйцеклетки видны темные точки, к которым подходят нити веретена деления, от которых радиально отходят волокна, образующие «звезду», или центросферу. Центросферу вместе с расположенными в центре центриоля ми называют центросомой. В животных клетках центриоли — центры организа ции микротрубочек веретена деления.
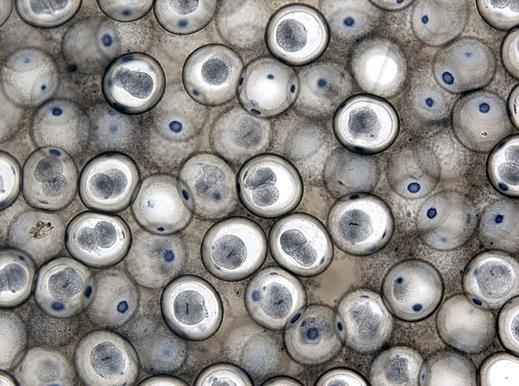
Митохондрии ( хондриосомы) в эпителиальных клетках кишечника аскариды.
Окраска: по Альтману
Сначала, используя малое увеличение микроскопа, необходимо найти базальный конец клеток кишечного эпителия, граничащий с соединительной тканью, и апикальный конец, направленный к просвету кишечника. На апикальном конце видна толстая кутикула. Ближе к базальному концу лежит овальное ядро.
Преимущественно в апикальной части клеток можно увидеть митохондрии, окрашенные в черный цвет. Они имеют вид коротких извитых нитей или палочек.
В эпителиальных клетках кишечника они поставляют энергию, заключенную в макроэргических связях АТФ, для осуществления функции всасывания.
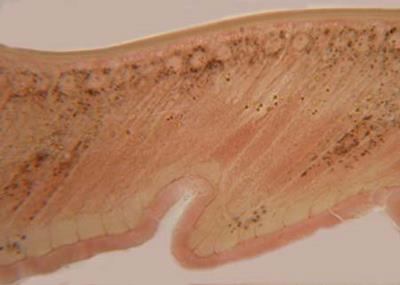

Аппарат Гольджи в нервных клетках спинального ганглия котенка.
Окраска: импрегнация серебром по методу К. Гольджи.
При слабом увеличении микроскопа выбрать нервную клетку, вокруг ядра которой хорошо заметна сетчатая структура аппарата Гольджи.. Обратить внимание на светлое круглое ядро нервной клетки. В центре ядра видно хорошо окрашенное ядрышко. Вокруг ядра в цитоплазме отметить аппарат Гольджи в виде черных глыбок, петель и изогнутых палочек. Аппарат, или комплекс Гольджи, в клетках выполняет функции модификации белков (связывает их с углеводами, фосфатными и сульфатными группами), формирования клеточной мембраны, образования лизосом.
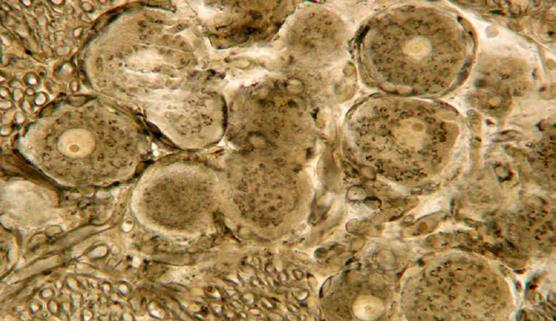 |
Секреторные гранулы в клетках Лейдига кожи аксолотля.
Окраска: гематоксилин-эозин.
Найти крупные (розового цвета) клетки Лейдига, лежащие близко к поверхностному кожному слою. Эти одноядерные клетки вырабатывают слизь – секрет белковой природы. Они — одноядерные, заполненные гранулами секрета. Зарисовать несколько клеток с гранулами секрета.
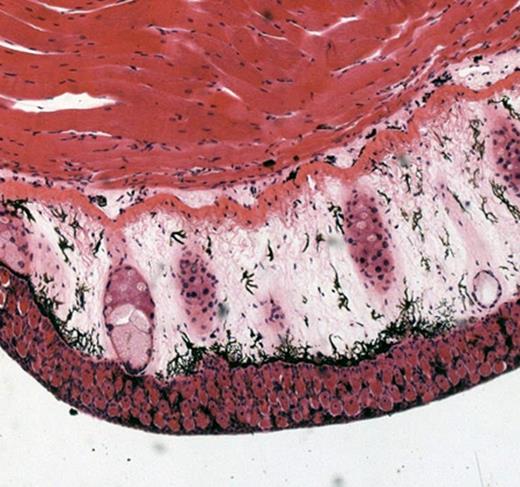
Желточные включения в бластомерах амфибии.
Окраска: гематоксилин-пикрофуксин.
Трофические белковые включения, то есть в качестве запаса питательных веществ, имеются только в яйцеклетках. Они содержатся в цитоплазме в виде гранул желтка. На препарате оплодотворенная яйцеклетка амфибии начинает дробиться на бластомеры. В их цитоплазме — золотисто-желтого цвета белковые гранулы. Рассмотреть на малом и большом увеличении микроскопа и зарисовать.

Включения гликогена в печени аксолотля.
Окраска: кармин Беста-гематоксилин.
При малом увеличении найдите, плотно расположенные многоугольные или округлые клетки гепатоцитов, окрашенные в ярко-розовый цвет. Ядро достаточно крупное, круглое и смещено к периферии, окрашено в фиолетовый цвет.
При большом увеличении видно, что содержимое каждой клетки представлено большим количеством глыбок гликогена. Глыбки имеют различную величину и в основном лежат ближе к одной из сторон клетки. Также среди глыбок гликогена и в той части цитоплазмы, где его нет, видны пустые неокрашенные вакуоли. В них находился жир и при приготовлении препарата (ксилол и спирт) он растворился.