


Электрические явления в нервах и мышцах
При определенных условиях в нервах и мышцах возникают электрические токи. Различают: 1) токи, или потенциалы, покоя и повреждения и 2) токи, или потенциал, действия – акционные точки.
Токи покоя
При отсутствии раздражения и, следовательно, возбуждения, существует разность потенциалов между наружной поверхностью цитоплазмы живой клетки (мембраной) и её внутренним содержимым.
Поверхностный слой цитоплазмы живой клетки, находиться в состоянии покоя, имеет положительный электрический заряд, а цитоплазма внутри клетки заряжена электроотрицательно. Поэтому, введя в клетку микроэлектрод и приложив другой электрод к поверхности клетки, можно отвести ток к регистрирующему прибору и измерить потенциал покоя, мембранный или клеточный потенциал.
Токи покоя, или клеточные потенциалы, обнаруживаются в неповрежденных живых клетках. Современными методами отведения электрического тока и его регистрации установлено, что токи покоя отдельных клеток животных, например гигантских нервных волокон кальмара, — 50-60 мв (милливольт), миелиновых нервных волокон амфибий – 70 мв, теплокровных – 90 мв, большинства тел нейронов – 80-100 мв, поперечнополосатых мышц амфибий – 80 мв, эпителия амфибий – 20 мв, волокон сердечной мышцы животных с постоянной температурой тела – 95 мв.
Токи повреждения
При повреждении живой ткани открывается доступ к внутреннему содержимому клеток, которое заряжено электроотрицательно. Поэтому если приложить электрод к неповрежденному участку ткани, находящемуся в состоянии покоя и, следовательно, заряженному электроположительно, а другой электрод приложить к поврежденному или перерожденному участку, заряженному электроотрицательно, то при соединении обоих электродов проводником тока с регистрирующим прибором можно измерить разность потенциалов.
Токи повреждения можно обнаружить на нервно-мышечном препарате. Если разрезать мышцу или любым способом повредить ее в одном участке (прижечь, раздавить и т. д.) и затем, приложив неполяризующиеся электроды, соединить срез мышцы или участок повреждения с целой поверхностью мышцы, не подвергающейся раздражению, то от целой поверхности к участку разреза или повреждения направится электрический ток. Сильно поврежденный участок всегда электроотрицателен по отношению к электроположительному неповрежденному участку. Следовательно, биоэлектрический ток повреждения, или альтерационный ток, возникает на границе целой и поврежденной частей ткани. Он обнаруживается и на некотором расстоянии от места повреждения, уменьшаясь с увеличением расстояния от места повреждения. Поэтому по месту происхождения его называют также демаркационным током. Однако ток повреждения появляется только тогда, когда отрицательно заряженный поврежденный участок соединяется проводником тока с положительно заряженным целым участком.
Величина тока повреждения меньше, чем клеточного потенциала, так как межклеточная и внеклеточная жидкости производят шунтирование (укорочение) при его измерении.
Разность потенциалов между целым и поврежденным участками скелетных мышц лягушки сразу же после повреждения равна 25-50 мв, наибольшая разность доходит до 80. В скелетных мышцах кролика токи повреждения равны 40 мв. В гладких мышцах токи повреждения меньше и быстрее уменьшаются до нуля. Разность потенциалов между целым и поврежденным участками седалищного нерва лягушки достигает 20-30 мв, седалищного нерва лошади — 6-16 мв, а седалищного нерва человека и обезьяны — 6-7 мв. Следовательно, токи повреждения нервов меньше, чем токи повреждения мышц. Особенно малы токи повреждения нервов человека и высших животных. Постепенно разность потенциалов уменьшается.
Когда нерв второго нервно-мышечного препарата набрасывается на первый нервно-мышечный препарат таким образом, чтобы он одновременно коснулся неповрежденного поврежденного участков мышцы, электрическая цепь замыкается. При этом мышца второго препарата сокращается в результате раздражения нерва током, возникающим в момент его замыкания между положительно заряженным нормальным участком мышцы первого препарата и отрицательно заряженным поврежденным ее участком.
Токи действия
Акционные токи, или токи (потенциалы) действия, возникают в раздражаемых тканях. Их появление характеризует возбуждение.
Для обнаружения тока действия нужно приложить один электрод к неповрежденному участку живой ткани, который находится в состоящих покоя и поэтому заряжен электроположительно, а другой — к раздражаемому участку и присоединить оба электрода к регистрирующему прибору. Раздражаемый участок при приложении к нему раздражителя немедленно или через несколько сотых миллисекунды становится электроотрицательным. Это падение потенциала’ доходит до максимума, а затем данный участок снова становится электроположительным. Различают однофазные и двухфазные токи действия.
Однофазный ток действия
При прикладывании одного из неполяризующихся электродов к неповрежденному участку нерва или мышцы, а другого — к поврежденному участку возникает электрический ток. Этот ток регистрируется при включении в цепь гальванометра или другого более чувствительною прибора.
Стрелка гальванометра отклонится в одну сторону вследствие электроотрицательного заряда поврежденного участка и электроположительного заряда нормального участка. Но если нанести раздражение по соседству с нормальным участком, то волна возбуждения, которая возникнет в месте раздражения, на некоторое время превратит заряд под нормальным участком в электроотрицательный. В этот момент тока не будет, так как под обоими электродами заряд станет электроотрицательным и стрелка гальванометра станет на нулевую линию. Когда волна возбуждения продвинется дальше и будет находиться между электродами, нормальный участок снова станет электроположительным и стрелка гальванометра снова отклонится в ту же сторону.
Когда волна возбуждения дойдет до поврежденного участка, положение стрелки гальванометра не изменится, так как под вторым электродом в месте повреждения уже имеется электроотрицательный заряд. Таким образом, стрелка гальванометра движется только в одну сторону, а запись этого движения регистрирует однофазный, или монофазный ток действия.
Это явление Дюбуа-Реймон назвал отрицательным колебанием тока покоя, которое возникает каждый раз, когда волна возбуждения проходит под электродом, расположенным на неповрежденном участке ткани.
Однофазный ток действия можно зарегистрировать также при раздражении ткани во время отведения клеточного потенциала. При внутриклеточном введении одного электрода и приложении другого электрода к поверхности клетки установлено, что клеточный потенциал не только исчезает в течение тысячных долей секунды, но что в течение этого промежутка времени амплитуда тока действия на 30-50 мв больше клеточного потенциала. Это объясняется тем, что в пункте прохождения волны возбуждения наружная поверхность мембраны становится электроотрицательной, а внутренняя — электроположительной. Продолжительность тока действия в нервных волокнах и клетках скелетных мышц составляет 0,1-5 мс.
Однофазный ток действия представляет собой местный электроотрицательный потенциал, мгновенно возникающий при очень слабых, подпороговых одиночных раздражениях и ограниченный несколькими миллиметрами от пункта раздражения. Величина местного низковольтного электроотрицательного заряда убывает по мере удаления от пункта раздражения. Чем больше подпороговое раздражение, тем больше возникающее возбуждение и амплитуда или отклонение однофазного тока действия.
Следовательно, местное возбуждение или местный сдвиг обмена веществ отличается градуальностью — возрастанием возбуждения по мере увеличения силы раздражения. Местное возбуждение наступает сразу после раздражения, т. е. без латентного периода. В участке слабого местного нераспространяющегося возбуждения возбудимость сохраняется, рефрактерность отсутствует. Если бы при местном возбуждении наблюдалась рефрактерность, то оно не могло бы перейти в пороговое, распространяющееся возбуждение.
Местный однофазный ток действия предшествует распространяющейся волне возбуждения и подготавливает ее. При одиночном пороговом раздражении волна возбуждения возрастает, достигает критического уровня и превращается из местной в распространяющуюся. Поэтому распространяющееся возбуждение наступает после латентного периода, продолжительность которого равна длительности местного нераспространяющегося возбуждения. В отличие от местного возбуждения распространяющееся возбуждение не градуально и сопровождается фазами изменения возбудимости. Потенциал действия, возникший в раздражаемом участке мембраны, вызывает возбуждение соседних участков мембраны и таким образом возбуждение распространяется по нервным и мышечным волокнам.
О прохождении волны возбуждения по возбудимой ткани судят прежде всего по токам действия. Однофазное колебание тока действия распространяющейся волны возбуждения в отдельных нервных и мышечных клетках состоит из нескольких составляющих: 1) высоковольтного потенциала, пика, 2) отрицательного следового потенциала и 3) положительного следового потенциала.
По сравнению со следовыми потенциалами пик имеет наибольшую высоту и наименьшую продолжительность. Пик имеет приблизительно одинаковую высоту на всем протяжении возбудимых тканей животных с постоянной температурой тела. Это указывает на то, что по мере распространения возбуждения вольтаж потенциала не падает. Этот факт свидетельствует, что химические вещества, необходимые для возникновения возбуждения, равномерно распределены по возбудимой ткани и что распространение возбуждения происходит самостоятельно, автоматически. Раздражение при достаточной интенсивности вызывает возбуждение только в том участке, который раздражается, а дальше возбуждение движется по ткани, вовлекая в сдвиг обмена веществ вещества, имеющиеся в возбудимой ткани.
Пик играет ведущую роль в распространении возбуждения. Высота пика различна у нервных волокон разных групп, имеющих неодинаковый диаметр, и изменяется в зависимости от физиологического состояния волокна и скорости проведения возбуждения. Чем толще нервное миелиновое волокно, тем меньше длительность пика. По мышечному волокну человека возбуждение проходит за 4-5 м/с.
В волокнах скелетных мышц однофазные токи действия различаются по амплитуде и по продолжительности. В волокнах сердечной мышцы они отличаются большей продолжительностью пика, который после начального крутого снижения начинает снижаться постепенно, а затем снова круто падает.
Токи действия в гигантских нервных волокнах кальмара достигают 90-110 мв, в миелиновых нервных волокнах амфибий 110 мв, в волокнах поперечнополосатых мышц амфибий — 120 мв, а в волокнах сердечной мышцы животных с постоянной темпера турой тела — 135 мв. Чем больше частота тока действия, тем меньше его вольтаж. Гладкие мышечные волокна отличаются тем, что их токи действия не превосходят мембранных потенциалов (не больше 70-80 мв).
Условия, улучшающие обмен веществ (повышение темпера туры, отдых), уменьшают продолжительность волны возбуждения, а условия, ухудшающие обмен веществ (утомление), увеличивают ее. С понижением температуры амплитуда пика уменьшается, а продолжительность его возрастает.
При температуре тела продолжительность подъема высоковольтного потенциала нерва составляет примерно 1/3 а продолжительность снижения — 2/3 его общей продолжительности. Предполагается, что крутой подъем пика обусловлен быстрым движением ионов натрия внутрь клетки.
Отрицательный следовой потенциал соответствует остаточным сдвигам распределения ионов калия и натрия. В отличие от пика он неустойчив и изменяется от условий среды. Он достигает 0,05 потенциала пика. Положительный низковольтный следовой потенциал также связан с перемещением ионов и равен примерно 0,002 потенциала пика.
Тепло и отдых уменьшают, а охлаждение и истощение увеличивают продолжительность следовой электроотрицательности. В нерве, лишенном кислорода, при действии наркоза и солей калия следовая низковольтная электроотрицательность отсутствует, а начальная высоковольтная электроотрицательность сохраняется, и при известных концентрациях наркоза даже не наступает заметных изменений её величины. Это указывает на то, что пик может возникать и при отсутствии кислорода, а следовая электроотрицательность связана с восстановительными процессами, которые совершаются в ткани вслед за возбуждением, и для её появления необходим кислород. Нарушение обмена веществ, утомление увеличивают продолжительность следовых потенциалов. Чем больше лабильность, тем меньше выражены следовые потенциалы. В высоколабильных мякотных волокнах человека и высших животных они почти не проявляются.
Электрические явления в нервах и мышцах развиваются в следующей последовательности. Сначала во все время раздражения отмечается электротоническое колебание, вызванное выходом катионов на поверхность клетки. Затем возникает местная низковольтная электроотрццательность. которая длится до начала появления высоковольтного потенциала. Это время от начала раздражения до начала пика, или латентный период, равно долям миллисекунды. После этого появляется пик; время»от начала его до вершины равно одной или нескольким миллисекундам. Продолжительность низковольтного отрицательного следового потенциала составляет несколько десятков миллисекунд и может доходить до 0,1 с и больше. Особенно велика продолжительность низковольтного положительного следового потенциала, которая равна десятым долям секунды.
Двухфазный ток действия
В естественных условиях одиночные волны возбуждения встречаются чрезвычайно редко; В организме в нервах и мышцах передаются серии волн возбуждения, взаимно влияющих друг на друга. Возбуждение, которое достигает порога или несколько превышает его, носит ритмический характер. В других случаях возбуждение в естественных условиях является градуальным, длительным, стойким, переходящим в торможение. В последнем случае при чрезмерно большой силе или частоте раздражения, превышающей меру лабильности, полностью угнетается высоковольтная электрическая активность.
Волновое распространяющееся возбуждение возникает из неволнового, градуального. Такая же эволюция возбуждения происходила в филогенезе. Представление о возникновении волнового возбуждения дает следующий простой опыт.
После приложения обоих неполяризующихся электродов к нормальным целым участкам возбудимой ткани и включения этой ткани в цепь гальванометра его стрелка останется на нуле, так как все участки нормальной ткани, находящейся в состоянии покоя, имеют одинаковый заряд. Но когда ткань раздражают у одного из электродов, то возникает возбуждение, и заряд ткани под этим электродом становится отрицательным. При этом стрелка гальванометра отклоняется в одну сторону. Передвигаясь далее, волна возбуждения оказывается между электродами, и в это время заряды под электродами вновь становятся одинаковыми и стрелка гальванометра возвращается к нулю. Но когда волна возбуждения доходит до второго электрода, то гальванометр опять отмечает разность потенциалов, так как под вторым электродом заряд становится отрицательным, а в то же самое время под первым электродом заряд положительный, так как под ним нег возбуждения. Теперь стрелка гальванометра отклоняется в противоположную сторону, а затем, после прекращения возбуждения, возвращается к нулю. Следовательно, когда по ткани проходит волна возбуждения, то стрелка гальванометра отклоняется сначала в одну сторону, потом в другую. Запись этого движения стрелки гальванометра регистрирует двухфазный ток действия.
Форма регистрации токов, или потенциалов, действия зависит от места возникновения возбуждения, от используемых приборов и от расположения электродов. В настоящее время их записывают посредством катодных осциллографов, не имеющих инерции, в отличие от гальванометров. Современные катодные осциллографы регистрируют биопотенциалы в миллионные доли вольта, продолжающиеся одну стомиллионную или даже миллиардную долю секунды. Эти токи отличаются сложностью, и их нередко расшифровывают при помощи электронных устройств.
Биопотенциалы каждой возбудимой ткани, например сердца, имеют характерный вид и изменяются в зависимости от ее функционального состояния. Записываются биопотенциалы кожи, эпителия, соединительной ткани, рецепторов, сетчатки глаза, скелетных мышц, пищеварительных желез, гладкой мускулатуры пищеварительного канала и других органов, разных отделов центральной нервной системы и даже одного нейрона. Первые исследования биопотенциалов больших полушарий головного мозга провели В. Я. Данилевский, Р. Кетон (1875), продолговатого мозга — И. М. Сеченов (1882).
Н. В. Введенский (1883, 1884) соединил мышцу проводами с телефонной трубкой и установил важнейшие законы, по которым протекает в ней возбуждение. Он же впервые применил телефон для изучения явлений возбуждения в нерве.
Презентация на тему: Введение в нормальную физиологию.
Физиология возбудимых тканей.
Лекция №1 Для медико-профилактического факультета
Кафедра нормальной физиологии им. Н.Ю.Беленкова доцент Продиус Петр Анатольевич
2013 г.
План лекции
•1. Предмет, задачи и методы исследований нормальной физиологии человека
•2. Понятие о возбудимых тканях и их свойства
•3. Современные представления о строении клеточной мембраны
•4. Мембранный потенциал и его происхождение
•5. Потенциал действия, его фазы и их происхождение.
Физиология как научная дисциплина
Физиология (греч. physis – природа, logos — учение) – наука, изучающая процессы жизнедеятельности и механизмы их регулирования в различных биологических системах: клетки – ткани – органы – системы органов – организм.
Предметом изучения физиологии являются функции живого организма, их связь между собой, регуляция и приспособление к внешней среде, происхождение и становление функций в процессе эволюции и индивидуального развития особи.
Физиологическая функция (functio-деятельность) – специфическая деятельность системы или органа, имеющая приспособительное значение и направленная на достижение полезного для организма результата.
Физиологический процесс – последовательность явлений в развитии какого-либо действия или совокупность последовательных действий, направленных на достижение определенного результата.

Живой организм как открытая биосистема
Система – совокупность элементов имеющих между собой связи.
Биосистема – совокупность живых клеток, тканей и органов постоянно взаимодействующих с внешней средой и осуществляющих с ней обмен веществ, энергии и информации.
Живые организмы более активно взаимодействуют с внешней средой чем объекты неживой природы
Основные понятия в физиологии возбудимых тканей
Раздражимость – способность живой материи активно отвечать на воздействие внешней и внутренней среды изменением обменных процессов.
Раздражитель – это изменение внешней или внутренней среды организма, воспринимаемое клетками и вызывающее ответную реакцию.
Возбудимость — способность ткани отвечать на раздражение быстрой деполяризацией мембраны, т.е. генерацией потенциала действия (ПД).
Возбуждение – процесс, характеризующийся изменением обмена клетки в ответ на раздражение в виде временной быстрой деполяризации мембраны, т.е. генерации ПД.
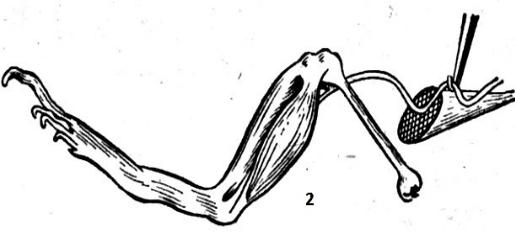
Ток покоя (повреждения)
•Открыт в 1794 г. во втором опыте Гальвани
– при набрасывании нерва на поврежденный и неповрежденный участок мышцы наблюдали мышечное сокращение .
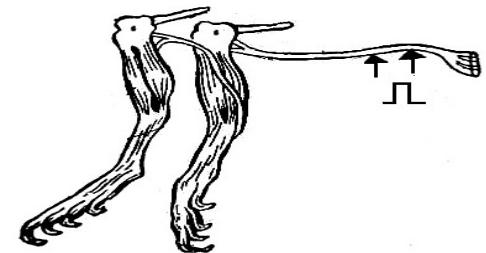
Ток действия
•Открыт в 1840 г. в опыте Маттеуччи (вторичный тетанус) – при раздражении индукционным током нерва первой реоскопической лапки наблюдали сокращение мышцы у первого и второго препаратов.
МЕМБРАННЫЙ ПОТЕНЦИАЛ ПОКОЯ
Дюбуа-Реймон: Поврежденный участок мембраны – заряжен «-», а неповрежденный – «+».
Бернштейн 1902 г. – расчет гипотетического мембранного потенциала по формуле Нернста.
Мембранно–ионная теория (Ходжкин, Хаксли, Катц (1949-1952).
Нобелевская премия в 1963 году.
Мембранный потенциал покоя (МПП) — разность
потенциалов между наружной и внутренней сторонами мембраны.
•Суть теории – мембранный потенциал покоя возникает благодаря направленному движению заряженных частиц.
•В основном это диффузия ионов К+ через мембрану клетки из внутриклеточной среды во внеклеточную.
•При создании мембранного потенциала покоя важную роль играют процессы простой диффузии через белковые каналы в мембране и первично активного транспорта.
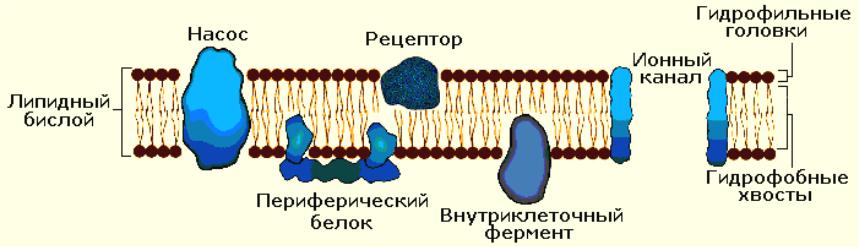
Строение мембраны
Жидкостно-мозаичная модель (С. Синджер и Дж.Николсон 1972 год ) – основу мембраны составляет двойной молекулярный слой липидов (липидный бислой) на котором и в толще которого находятся белки. Слой липидов является прерывистым, белки клеточной мембраны подвижны и свободно плавают в липидном геле.
Избирательная проницаемость клеточной мембраны
Обмен веществ через мембрану клетки осуществляется через липидный бислой или транспортные белки с помощью процессов диффузии, осмоса, активного и везикулярного транспорта.
Простая диффузия – пассивное движение частиц из области высокой концентрации в область низкой концентрации. Жирорастворимые вещества диффундируют через липидный бислой (этанол, СО2),
водорастворимые через водные каналы (ионы К+, Na+). Ионные каналы делятся на управляемые и неуправляемые.
Осмос – пассивное движение воды из области с низкой концентрацией веществ в область с высокой концентрацией веществ.
Общая физиология возбудимых тканей

1. Мембраны в основном состоят из липидов и белков, количество которых неодинаково у разных типов клеток. Сложность изучения молекулярных механизмов функционирования клеточных мембран обусловлена тем, что при выделении и очистке клеточных мембран нарушается их нормальное функционирование. В настоящее время можно говорить о нескольких видах моделей клеточной мембраны, среди которых наибольшее распространение получила жидкостно-мозаичная модель.Согласно этой модели, мембрана представлена бислоем фосфолипидных молекул, ориентированных таким образом, что гидрофобные концы молекул находятся внутри бислоя, а гидрофильные направлены в водную фазу (рис. 2.1). Такая структура идеально подходит для образования раздела двух фаз: вне- и внутриклеточной. В фосфолипидном бислое интегрированы глобулярные белки, полярные участки которых образуют гидрофильную поверхность в водной фазе. Эти интегрированные белки выполняют различные функции, в том числе рецепторную, ферментативную, образуют ионные каналы, являются мембранными насосами и переносчиками ионов и молекул. Некоторые белковые молекулы свободно диффундируют в плоскости липидного слоя; в обычном состоянии части белковых молекул, выходящие по разные стороны клеточной мембраны, не изменяют своего положения. Здесь описана только общая схема строения клеточной мембраны и для других типов клеточных мембран возможны значительные различия. Электрические характеристики мембран. Особая морфология клеточных мембран определяет их электрические характеристики, среди которых наиболее важными являются емкость и проводимость. Емкостные свойства в основном определяются фосфолипидным бислоем, который непроницаем для гидратированных ионов и в то же время достаточно тонок (около 5 нм), чтобы обеспечивать эффективное разделение и накопление зарядов и электростатическое взаимодействие катионов и анионов. Кроме того, емкостные свойства клеточных мембран являются одной из причин, определяющих временные характеристики электрических процессов, протекающихщих на клеточных мембранах. Проводимость (g) — величина, обратная электрическому сопротивлению и равная отношению величины общего трансмембранного тока для данного иона к величине, обусловившей его трансмембранной разности потенциалов. Через фосфолипидный бислой могут диффундировать различные вещества, причем степень проницаемости (Р), т. е. способность клеточной мембраны пропускать эти вещества, зависит от разности концентраций диффундирующего вещества по обе стороны мембраны, его растворимости в липидах и свойств клеточной мембраны. Скорость диффузии для заряженных ионов в условиях постоянного поля в мембране определяется подвижностью ионов, толщиной мембраны, распределением ионов в мембране. Для неэлектролитов проницаемость мембраны не влияет на ее проводимость, поскольку неэлектролиты не несут зарядов, т. е. не могут переносить электрический ток. Проводимость мембраны является мерой ее ионной проницаемости. Увеличение проводимости свидетельствует об увеличении количества ионов, проходящих через мембрану.
ФУНКЦИИ:
1.Барьерная функция выражается в том, что мембрана при помощи соответствующих механизмов участвует в создании концентрационных градиентов, препятствуя свободной диффузии. При этом мембрана принимает участие в механизмах электрогенеза. К ним относятся механизмы создания потенциала покоя, генерация потенциала действия, механизмы распространения биоэлектрических импульсов по однородной и неоднородной возбудимым структурам.
2.Регуляторная функция клеточной мембраны заключается в тонкой регуляции внутриклеточного содержимого и внутриклеточных реакций за счет рецепции внеклеточных биологически активных веществ, что приводит к изменению активности ферментных систем мембраны и запуску механизмов вторичных «месенджеров» («посредников»).
3.Преобразование внешних стимулов неэлектрической природы в электрические сигналы (в рецепторах).
4.Высвобождение нейромедиаторов в синаптических окончаниях.
5.На ее поверхности протекает большинство биохимических реакций.
6.Мембрана координирует и регулируют физические процессы в клетке.
7.мембрана играет важную роль в образовании межклеточных контактов.
По способу активации выделяют:
потенциал-активируемые ионные каналы (переход из закрытого в открытое состояние и обратно осуществляется конформацией белковой молекулы при изменении потенциала мембраны). Примером может служить потенциал-зависимый натриевый канал, определяющий деполяризацию клетки при генерации потенциала действия.
механочувствительные ионные каналы (открываются при воздействии на мембрану клетки механического стимула, например, при активации механорецепторов кожи).
лиганд-активируемые ионные каналы. По способу активации они подразделены на две группы (экстраклеточные и внутриклеточные) в зависимости от того, с какой стороны мембраны воздействует лиганд. Если стимул (например, ацетилхолин) при осуществлении синаптической передачи возбуждения в нервно-мышечном синапсе действует на рецептор (в данном примере холинорецептор, представляющий собой одну из нескольких белковых субъединиц ионного канала), расположенный на внешней поверхности мембраны мышечной клетки, откроется ионный канал, проницаемый для катионов. Если лиганд-активируемые каналы зависят от вторичных посредников в клетке, их переход в открытое состояние осуществляется при изменении концентрации определенных ионов в цитоплазме. Примером может служить кальций-активируемый калиевый канал, активирующийся при увеличении концентрации ионов кальция в клетке. Такие каналы принимают участие в реполяризации мембраны при завершении потенциала действия.
Селективность, или избирательность, канала обеспечивается его особой белковой структурой. Большинство каналов являются электроуправляемыми, т. е. их способность проводить ионы зависит от величины мембранного потенциала. В состоянии покоя натриевый канал закрыт. При деполяризации клеточной мембраны до определенного уровня происходит открытие m-активационных ворот (активация) и усиление поступления ионов Na+ внутрь клетки. Через несколько миллисекунд после открытия m-ворот происходит закрытие п-ворот, расположенных у выхода натриевых каналов (инактивация) (рис. 2.4). Инактивация развивается в клеточной мембране очень быстро и степень инактивации зависит от величины и времени действия деполяризующего стимула.
Кроме натриевых, в клеточных мембранах установлены другие виды каналов, избирательно проницаемых для отдельных ионов: К+, Са2+, причем существуют разновидности каналов для этих ионов (см. табл. 2.1).
Свойство проводимости различных каналов неодинаково. В частности, для калиевых каналов процесс инактивации, как для натриевых каналов, не существует. Имеются особые калиевые каналы, активирующиеся при повышении внутриклеточной концентрации кальция и деполяризации клеточной мембраны. Активация калий-кальцийзависимых каналов ускоряет реполяризацию, тем самым восстанавливая исходное значение потенциала покоя.
Особый интерес представляют кальциевые каналы.
Входящий кальциевый ток, как правило, недостаточно велик, чтобы нормально деполяризовать клеточную мембрану. Чаще всего поступающий в клетку кальций выступает в роли «мессенджера», или вторичного посредника. Активация кальциевых каналов обеспечивается деполяризацией клеточной мембраны, например входящим натриевым током.
Процесс инактивации кальциевых каналов достаточно сложен. С одной стороны, повышение внутриклеточной концентрации свободного кальция приводит к инактивации кальциевых каналов. С другой стороны, белки цитоплазмы клеток связывают кальций, что позволяет поддерживать длительное время стабильную величину кальциевого тока, хотя и на низком уровне; при этом натриевый ток полностью подавляется. Кальциевые каналы играют существенную роль в клетках сердца.
Электровозбудиые: раздражитель (электрический ток) -> сдвиг мембранного потенциала(до критического потенциала) -> активация потенциалуправляемых ионных каналов -> изменение ионной проницаемости мембраны -> изменение ионных токов через мембрану -> дальнейший сдвиг мембранного потенциала( ответ в виде формирования потенциала действия)
Хемовозбудимые: раздражитель( химическое вещество) -> химическое связывание раздражителя и рецептора хемоуправляемого ионного канала -> изменение конформации лигандрецепторного комплекса и открытие рецептруправляемых( хемоуправляемых) ионных каналов-> изменение ионной проницаемости мембраны -> изменение ионных токов через мембрану -> дальнейший сдвиг мембранного потенциала( ответ в виде формирования потенциала действия)
Механовозбудимые: раздражитель (механический стресс)-> изменение натяжения мембраны -> открытие механоуправляемых ионных каналов -> -> изменение ионной проницаемости мембраны -> изменение ионных токов через мембрану -> дальнейший сдвиг мембранного потенциала( ответ в виде формирования потенциала действия)
2. Все клетки имеют свой электрический заряд, который формируется в результате неодинаковой проницаемости мембраны для различных ионов. Клетки возбудимых тканей (нервная, мышечная, железистая) отличаются тем, что они под действием раздражителя меняют проницаемость своей мембраны для ионов, в результате чего ионы очень быстро транспортируются согласно электрохимическому градиенту. Это и есть процесс возбуждения. Его основой является потенциал покоя. Потенциал покоя – относительно стабильная разность электрических потенциалов между наружной и внутренней сторонами клеточной мембраны. Его величина обычно варьирует в пределах от -30 до -90 мВ. Внутренняя сторона мембраны в покое заряжена отрицательно, а наружная – положительно из-за неодинаковых концентраций катионов и анионов внутри и вне клетки.
основную роль в создании отрицательного заряда внутри клетки играют ионы K+ и высокомолекулярные внутриклеточные анионы, главным образом они представлены белковыми молекулами с отрицательно заряженными аминокислотами (глутамат, аспартат) и органическими фосфатами. Эти анионы, как правило, не могут транспортироваться через мембрану, создавая постоянный отрицательный внутриклеточный заряд. Во всех точках клетки отрицательный заряд практически одинаков. Заряд внутри клетки является отрицательным как абсолютно (в цитоплазме анионов больше, чем катионов), так и относительно наружной поверхности клеточной мембраны. Абсолютная разность невелика, однако этого достаточно для создания электрического градиента. Обеспечивающим формирование потенциала покоя (ПП), является K+. В покоящейся клетке устанавливается динамическое равновесие между числом входящих и выходящих ионов K+. Это равновесие устанавливается тогда, когда электрический градиент уравновесит концентрационный. Согласно концентрационному градиенту, создаваемому ионными насосами, K+ стремится выйти из клетки, однако отрицательный заряд внутри клетки и положительный заряд наружной поверхности клеточной мембраны препятствуют этому (электрический градиент). В случае равновесия на клеточной мембране устанавливается равновесный калиевый потенциал. В покое вход Na+ в клетку низкий (намного ниже, чем K+), но он уменьшает мембранный потенциал. Влияние Cl- противоположно, так как это анион. Отрицательный внутриклеточный заряд не позволяет большому количеству Cl- проникнуть в клетку, поэтому Cl- это в основном внеклеточный анион. Как внутри клетки, так и вне ее Na+ и Cl- нейтрализуют друг друга, вследствие чего их совместное поступление в клетку не оказывает существенного влияния на величину потенциала покоя.
Наружная и внутренняя стороны мембраны несут на себе собственные электрические заряды, преимущественно с отрицательным знаком. Это полярные составляющие мембранных молекул – гликолипидов, фосфолипидов, гликопротеинов. Ca2+, как внеклеточный катион, взаимодействует с наружными фиксированными отрицательными зарядами, а также с отрицательными карбоксильными группами интерстиция, нейтрализуя их, что приводит к увеличению и стабилизации потенциала покоя.
Для создания и поддержания электрохимических градиентов необходима постоянная работа ионных насосов. Ионный насос – это транспортная система, обеспечивающая перенос иона вопреки электрохимическому градиенту, с непосредственными затратами энергии. Градиенты Na+ и K+ поддерживаются с помощью Na/K – насоса. Сопряженность транспорта Na+ и K+ примерно в 2 раза уменьшает энергозатраты. В целом же траты энергии на активный транспорт огромны: лишь Na/K – насос потребляет около 1/3 всей энергии, расходуемой организмом в покое. 1АТФ обеспечивает один цикл работы – перенос 3 Na+ из клетки, и 2 K+ в клетку. Асимметричный перенос ионов способствует заодно формированию и электрического градиента (примерно 5 – 10мВ).
Активный транспорт- с затратой энергии АТФ, против концентрационного и электрического градиента(насосы)
-первичный(включает перенос отдельных ионов вопреки концентрационному и электрическому градиентам с помощью специальных ионных насосов, микровезикулярный транспорт( эндоцитоз, экзоцитоз, трансцитоз)и фильтрацию.
-вторичный(он осуществляется за счет энергии, запасенной ранее, которая создается в виде электрического и концентрационного градиентов, т. е. тоже в результате расхода энергии, поэтому называть его пассивным необоснованно, хотя частицы движутся согласно законам диффузии.)
Пассивный транспорт- без затраты энергии по концентрационному и электрическому градиенту.
-простая диффузия(ионные каналы)
-облегченная диффузия(белки-переносчики)
-осмос
В результате непрерывного перемещения различных ионов через клеточную мембрану их концентрация внутри и вне клетки постепенно должна выравниваться. Имеется активный механизм поддержания градиентов концентрации различных ионов внутри и вне клетки. Им являются ионные насосы, в частности Na/K- насос.
Ионный насос — обладающая АТФ-азной активностью молекула интегрального белка, обеспечивающая перенос ионов через мембрану с непосредственной затратой энергии вопреки концентрационному и электрическому градиентам.выведение Na+ сопряжено с транспортом К+, что мжно продемонстрировать при адалении К+ из наружного раствора. Если К+ вне клетки нет, работа насоса блокируется, перенос Na+ из клетки в этом случае падает,составляя примерно 30% от нормально уровня. Накопление Na+ в клетке стимулирует работу насоса, уменьшение Na+ в клетке снижает его активность, поскольку снижается вероятность контакта ионов с соответствующим переносчиком. В результате сопряженного транспорта Na+ K+ поддерживается постоянная разность концентраций этих ионов внутри и вне клетки. Одна молекула АТФ- обеспечивает один цикл работы Na/K насоса: перенос трех ионов Na+ за пределы клетки и двух ионов К+ внутрь клетки. Такой ассиметричный перенос ионов поддерживает избыток положительно заряженных частиц вне клетки и отрицательных зарядов внутри клетки,что позволяет считать Na/K насос структурной электрогенной,дополнительно увеличивающей потенциал покоя примерно на несколько милливольт.
Блокаторы — это вещества, препятствующее работе ионного канала, например, взаимодействию медиатора с молекулярным рецептором к нему и, следовательно, нарушающие управление каналом, блокирующие его. Например, действие ацетилхолина блокируют холиноблокаторы; норадреналина с адреналином — адреноблокаторы; гистамина — гистаминоблокаторы и т. д. Многие блокаторы применяются в терапевтических целях как лекарственные препараты.
Блокаторы — это вещества, препятствующее работе ионного канала, например, взаимодействию медиатора с молекулярным рецептором к нему и, следовательно, нарушающие управление каналом, блокирующие его. Например, действие ацетилхолина блокируют холиноблокаторы; норадреналина с адреналином — адреноблокаторы; гистамина — гистаминоблокаторы и т. д. Многие блокаторы применяются в терапевтических целях как лекарственные препараты.
3.
Два основных класса рецепторов — это метаботропные рецепторыиионотропные рецепторы.
Ионотропные рецепторы представляют собой мембранные каналы, открываемые или закрываемые при связывании с лигандом. Возникающие при этом ионные токи вызывают изменения трансмембранной разности потенциалов и, вследствие этого, возбудимости клетки, а так же меняют внутриклеточные концентрации ионов, что может вторично приводитъ к активации систем внутриклеточных посредников. Одним из наиболее полно изученных ионотропных рецепторов являетсян-холинорецептор.
Метаботропные рецепторы связаны с системами внутриклеточных посредников. Изменения их конформации при связывании с лигандом приводит к запуску каскада биохимических реакций, и, в конечном счете, измемению функционального состояния клетки.
Рецепторы, связанные с гетеротримерными G-белками(например, рецепторвазопрессина).
Рецепторы, обладающие внутренней тирозинкиназной активностью (например, рецепторинсулина).
Внутриклеточные рецепторы-факторы транскрипции (например, рецепторы глюкокортикоидов).
Рецепторы, связанные с G-белками, представляют собой трансмембранные белки, имеющие 7 трансмембранных доменов, внеклеточный N-конец и внутриклеточный C-конец. Сайт связывания с лигандом находится на внеклеточных петлях, домен связывания с G-белком — вблизи C-конца в цитоплазме.
Активация рецептора приводит к тому, что его α-субъединицадиссоциирует от βγ-субъединичного комплекса и таким образом активируется. После этого она либо активирует, либо наоборот инактивируетфермент, продуцирующий вторичные посредники.
Рецепторы с тирозинкиназной активностью фосфорилируютпоследующие внутриклеточные белки, часто тоже являющиеся протеинкиназами, и таким образом передают сигнал внутрь клетки. По структуре это — трансмембранные белки с одним мембранным доменом. Как правило, гомодимеры, субъединицы которых связаныдисульфидными мостиками. Внутриклеточные рецепторы после связывания с гормоном переходят в активное состояние, транспортируются в ядро клетки, там связываются с ДНК и либо индуцируют, либо супрессируют экспрессию некоторого гена или группы генов.
Имеется несколько видов G-белков (GS, Gi ,Gq ,GO), которые влияют на ферменты (например, аденилатциклазу, фосфолипазу С), образующие различные вторые посредники (например, цАМФ, инозитолтрифосфат, диацилглицерол). При этом G-белки могут как активировать (GS), так и ингибировать (Gi) образование вторых посредников. Вторые посредники, в свою очередь, активируют соответствующие им протеинкиназы, фосфорилирующие различные клеточные белки по остаткам серина и треонина, что изменяет их функциональную активность (чаще увеличивает). Наряду с этим активированные G-белки оказывают выраженное влияние на проницаемость кальциевых и калиевых каналов мембраны.
4 вопрос Мембранный потенциал (или потенциал покоя) – это разность потенциалов между наружной и внутренней поверхностью мембраны в состоянии относительного физиологического покоя. Потенциал покоя возникает в результате двух причин: 1) неодинакового распределения ионов по обе стороны мембраны; 2) избирательной проницаемости мембраны для ионов. В состоянии покоя мембрана неодинаково проницаема для различных ионов. Клеточная мембрана проницаема для ионов K, малопроницаема для ионов Na и непроницаема для органических веществ. За счет этих двух факторов создаются условия для движения ионов. Это движение осуществляется без затрат энергии путем пассивного транспорта – диффузией в результате разности концент-рации ионов. Ионы K выходят из клетки и увеличивают положительный заряд на наружной поверхности мембраны, ионы Cl пассивно переходят внутрь клетки, что приводит к увеличению положительного заряда на наружной поверхности клетки. Ионы Na накапливаются на наружной поверхности мембраны и увеличивают ее положительный заряд. Органические соединения остаются внутри клетки. В результате такого движения наружная поверхность мембраны заряжается положительно, а внутренняя – отрицательно. Внутренняя поверхность мембраны может не быть абсолютно отрицательно заряженной, но она всегда заряжена отрицательно по отношению к внешней. Такое состояние клеточной мембраны называется состоянием поляризации. Движение ионов продолжается до тех пор, пока не уравновесится разность потенциалов на мембране, т. е. не наступит электрохимическое равновесие. Момент равновесия зависит от двух сил: 1) силы диффузии; 2) силы электростатического взаимодействия. Значение электрохимического равновесия: 1) поддержание ионной асимметрии; 2) поддержание величины мембранного потенциала на постоянном уровне. В возникновении мембранного потенциала участвуют сила диффузии (разность концентрации ионов) и сила электростатического взаимодействия, поэтому мембранный потенциал называется концентра-ционно-электрохимическим. Для поддержания ионной асимметрии электрохимического равновесия недостаточно. В клетке имеется другой механизм – натрий-калиевый насос. Натрий-калиевый насос – механизм обеспечения активного транспорта ионов. В клеточной мембране имеется система переносчиков, каждый из которых связывает три иона Na, которые находятся внутри клетки, и выводит их наружу. С наружной стороны переносчик связывается с двумя ионами K, находящимися вне клетки, и переносит их в цитоплазму. Энергия берется при расщеплении АТФ. Любая живая клетка поддерживает на мембране определенной величины потенциал. Его величина колеблется в значительных пределах, у нервных и мышечных клеток его величина составляет обычно 60 — 90 мВ, у других тканей не превышает 10 мВ. Различные внешние воздействия, способные изменяющие ионную проницаемость мембраны (то есть повлиять на воротный механизм канала), вызывают изменения величины ПП. Его уменьшение называется деполяризацией, а увеличение — гиперполяризацией мембраны
5 вопрос Возбужде́ние в физиологии — ответ ткани на раздражение, проявляющийся помимо неспецифических реакций (генерация потенциала действия, метаболические изменения) в выполнении специфической для этой ткани функции; возбудимыми являются нервная (проведение возбуждения), мышечная (сокращение) и железистая (секреция) ткани. Возбудимость — свойство клеток отвечать на раздражение возбуждением. Реакция клеток, тканей на раздражитель определяется законами раздражения 1. Закон «все или ничего»: При допороговых раздражениях клетки в ткани ответной реакции не возникает. При пороговой силе раздражителя развивается максимальная ответная реакция, поэтому увеличение силы раздражения выше пороговой не сопровождается ее усилением. В соответствии с этим законом реагирует на раздражения одиночное нервное и мышечное волокно, сердечная мышца. 2. Закон силы: Чем больше сила раздражителя, тем сильнее ответная реакция. Однако выраженность ответной реакции растет лишь до определенного максимума. Закону силы подчиняется целостная скелетная, гладкая мышца, так как они состоят из многочисленных мышечных клеток, имеющих различную возбудимость. 3. Закон силы-длительности. Между силой и длительностью действия раздражителя имеется определенная взаимосвязь. Чем сильнее раздражитель, тем меньшее время требуется для возникновения ответной реакции. Зависимость между пороговой силой и необходимой длительностью раздражения отражается кривой силы длительности. По этой кривой можно определить ряд параметров возбудимости. а) Порог раздражения — это минимальная сила раздражителя, при которой возникает возбуждение. б) Реобаза — это минимальная сила раздражителя, вызывающая возбуждение при его действии в течение неограниченно долгого времени. На практике порог и реобаза имеют одинаковый смысл. Чем ниже порог раздражения или меньше реобаза, тем выше возбудимость ткани. в) Полезное время — минимальное время действия раздражителя силой в одну реобазу за которое возникает возбуждение. г) Хронаксия — это минимальное время действия раздражителя силой в две реобазы, необходимое для возникнов4. Закон градиента или аккомодации. Реакция ткани на раздражение зависит от его градиента, т.е. чем быстрее нарастает сила раздражителя во времени тем быстрее возникает ответная реакция. При низкой скорости нарастания силы раздражителя растет порог раздражения. Поэтому если сила раздражителя , возрастает очень медленно возбуждения не будет. Это явление называется аккомодацией. Физиологическая лабильность (подвижность) — это большая или меньшая частота реакций, которыми может отвечать ткань на ритмическое раздражение. Чем быстрее восстанавливается ее возбудимость после очередного раздражения, тем Выше ее лабильность. Определение лабильности предложено Н.Е.Введенским. Наибольшая, лабильность у нервов, наименьшая у сердечной мышцы. ения возбуждения. Действие постоянного тока на возбудимые ткани. •возбудимость повышается, а под положительным — анодом снижается. Это называется законом действия постоянного Тока. Изменение возбудимости ткани (например: нерва) под действием постоянного тока в области анода или катода называется физиологическим электротоном. В настоящее время установлено, что под действием отрицательного электрода — катода потенциал мембраны клеток снижается. Это явление называется физическим катэлектротоном, Под положительным — анодом, он возрастает. Возникает физический катэлектртон. Так как, под катодом мембранный потенциал приближается к критическому уровню деполяризации, возбудимость клеток и тканей повышается. Под анодом мембранный потенциал возрастает и удаляется от критического уровня деполяризации, поэтому возбудимость клетки, ткани падает. Следует отметить, что при очень кратковременном действии постоянного тока (1 мсек и менее)МП не успевает измениться, поэтому не изменяется и возбудимость ткани под электродами. Критический уровень деполяризации (Ек)- это та разность потенциалов, которая должна быть достигнута, чтобы местные изменения перешли в пик потенциала действия. Ек — пороговая величин
6 вопрос Фаза деполяризации возникает в результате лавинообразного движения Nа+ внутрь клетки. Этому способствуют две причины: открываются потенциалзависимые Nа+-каналы. В этом случае происходит деполяризация по типу процесса с положительной обратной связью (самоподкрепляющийся процесс). Освобождение натриевых каналов от Са2+. Заряд клеточной мембраны сначала снижается до 0 (это собственно деполяризация), а затем меняется на противоположный (инверсия или овершут). Для характеристики фазы деполяризации вводится понятие реверсии — это та разность потенциалов, на которую потенциал действия превышает потенциал покоя. Р (реверсия)- это то количество мВ на которое произошла перезарядка мембраны. Фаза деполяризации продолжается до достижения электрохимического равновесия по Nа+. Затем наступает следующая фаза. Амплитуда потенциала действия не зависит от силы раздражителя. Она зависит от концентрации Nа+ (как снаружи, так и внутри клетки), от количества натриевых каналов, особенностей натриевой проницаемости. Фаза реполяризации характеризуется: снижением проницаемости клеточной мембраны для Nа+ (Nа-инактивация). Натрий накапливается на наружной поверхности клеточной мембраны; возрастает проницаемость мембраны для К+, в результате повышается выход К+ из клетки с увеличением положительного заряда на мембране; изменение активности Nа+-К+ насоса. Реполяризация — это процесс восстановления заряда мембраны. Но полного восстановления нет, т. к. возникают следовые потенциалы. Гиперполяризация – при частом возбуждении канал не успевает захлопнуться, калий выходит из клетки, мембрана становится более электроотрицателной. Так как в формировании потенциала покоя мембраны участвуют многие ионы, равновесие клетки может нарушаться посредством изменений проводимости различных ионов. Так, например, при дополнительном выходящем токе ионов К+ или при входящем токе ионов Сl может увеличиваться потенциал покоя мембраны, это означает что она гиперполяризуется. Гиперполяризация мембраны — противоположность возбуждения, т.е. определенные химические процессы на постсинаптической мембране могут вызывать торможение нейрона.
7. Потенциал действия – это сдвиг мембранного потенциала, возникающий в ткани при действии порогового и сверхпорогового раздражителя, что сопровождается перезарядкой клеточной мембраны.
При действии порогового или сверхпорогового раздражителя изменяется проницаемость клеточной мембраны для ионов в различной степени. Для ионов Na она повышается в 400–500 раз, и градиент нарастает быстро, для ионов К – в 10–15 раз, и градиент развивается медленно. В результате движение ионов Na происходит внутрь клетки, ионы К двигаются из клетки, что приводит к перезарядке клеточной мембраны. Наружная поверхность мембраны несет отрицательный заряд, внутренняя – положительный.
Стадия покоя. Эта стадия представлена мембранным потенциалом покоя, который предшествует потенциалу действия. Мембрана во время этой стадии поляризована в связи с наличием отрицательного мембранного потенциала
Фаза деполяризации. В это время мембрана внезапно становится высокопроницаемой для ионов натрия, позволяя огромному числу положительно заряженных ионов натрия диффундировать внутрь клетки. Нормальное поляризованное состояние немедленно нейтрализуется поступающими внутрь положительно заряженными ионами натрия, в результате потенциал стремительно нарастает в положительном направлении. Этот процесс называют деполяризацией,
Фаза реполяризации. В течение нескольких долей миллисекунды после резкого повышения проницаемости мембраны для ионов натрия, натриевые каналы начинают закрываться, а калиевые — открываться. В результате быстрая диффузия ионов калия наружу восстанавливает нормальный отрицательный мембранный потенциал покоя. Этот процесс называют реполя-ризацией мембраны.
Изменения мембранного потенциала,следующие за пиком потэнциала действия,называют СЛЕДОВЫМИ ПОТЕНЦИАЛАМИ.
Различают 2 вида следовых потенциалов-следовую деполяризацию и следовую гиперполяризацию.Амплитуда следовых потенциалов обычно не превышает нескольких милливольт,а длительность их у различных волокон составляет от нескольких миллисекунд до десятков и сотен секунд.
Механизм проведения возбуждения по нервным волокнам зависит от их типа. Существуют два типа нервных волокон: миелиновые и безмиелиновые.
Процессы метаболизма в безмиелиновых волокнах не обеспечивают быструю компенсацию расхода энергии. Распространение возбуждения будет идти с постепенным затуханием – с декрементом. Декрементное поведение возбуждения характерно для низкоорганизованной нервной системы. Возбуждение распространяется за счет малых круговых токов, которые возникают внутрь волокна или в окружающую его жидкость. Между возбужденными и невозбужденными участками возникает разность потенциалов, которая способствует возникновению круговых токов. Ток будет распространяться от «+» заряда к «—». В месте выхода кругового тока повышается проницаемость плазматической мембраны для ионов Na, в результате чего происходит деполяризация мембраны. Между вновь возбужденным участком и соседним невозбужденным вновь возникает разность потенциалов, что приводит к возникновению круговых токов. Возбуждение постепенно охватывает соседние участки осевого цилиндра и так распространяется до конца аксона.
В миелиновых волокнах благодаря совершенству метаболизма возбуждение проходит, не затухая, без декремента. За счет большого радиуса нервного волокна, обусловленного миелиновой оболочкой, электрический ток может входить и выходить из волокна только в области перехвата. При нанесения раздражения возникает деполяризация в области перехвата А, соседний перехват В в это время поляризован. Между перехватами возникает разность потенциалов, и появляются круговые токи. За счет круговых токов возбуждаются другие перехваты, при этом возбуждение распространяется сальтаторно, скачкообразно от одного перехвата к другому. Сальтаторный способ распространения возбуждения экономичен, и скорость распространения возбуждения гораздо выше (70—120 м/с), чем по безмиелиновым нервным волокнам (0,5–2 м/с).
Возникновению потенциала действия предшествует в точке раздражения мышцы или нерва активные под пороговые изменения мембранного потенциала. Они проявляются в форме локального (местного) ответа.
8. Для локального ответа характерны:
1) зависимость от силы раздражения
2) нарастание постепенно величины ответа.
3) нераспространение по нервному волокну.
Первые признаки локального ответа обнаруживаются при действии стимулов составляющих 50-70% пороговой величины. Локальный ответ как и потенциал действия обусловлен повышением натриевой проницаемости. Однако это повышение было недостаточно, чтобы вызвать потенциал действия.
Потенциал действия возникает когда деполяризация мембраны достигнет критического уровня. Но локальный ответ важен. Он подготавливает ткани к последующим воздействиям.
Сравнительная характеристика локального потенциала и ПД
Свойство | Локальный потенциал | Потенциал действия |
Распространение | На 1 – 2 мм с затуханием (декрементом) | Без декремента на большие расстояния по всей длине нервного волокна |
Зависимость от величины стимула | Возрастает с увеличением силы раздражителя, т. е. подчиняется закону «силы» | Не зависит (подчиняется закону «все или ничего») |
Явление суммации | Суммируется – возрастает при частых повторных подпороговых раздражениях | Не суммируется |
Амплитуда | 10 – 40 мВ | 80 – 130 мВ |
Возбудимость ткани при возникновении потенцала | Увеличивается | Уменьшается вплоть до абсолютной рефрактерности |
Повышение возбудимости клетки во время локального потенциала объясняется тем, что мембрана оказывается частично деполяризованной. Если КУД остается на постоянном уровне, то для его достижения требуется гораздо меньший раздражитель. Амплитуда ПД не зависит от силы раздражителя, потому что он возникает вследствие регенеративных процессов.
При стимуляции рецептора внешним воздействием, к которому чувствителен рецептор , происходит деполяризация сомы , которая исчезает с прекращением стимуляции. Деполяризация называется рецепторным потенциалом (или генераторным потенциалом ). Длительность его соответствует длительности стимула, а его амплитуда возрастает с увеличением интенсивности стимуляции, таким образом, он является отражением стимула, а не ответом по типу «все или ничего», как потенциал действия . Рецепторный потенциал обусловлен повышением Na+ — проводимости мембраны дендритов , в результате чего вход ионов натрия создает деполяризующий рецепторный потенциал, который электротонически распространяется к соме . Эта первичная трансформация стимула в рецепторный потенциал называется преобразованием, а рецептор, таким образом, является преобразователем, датчиком.
Пороговая Сила тока увеличивается при уменьшении крутизны его нарастания,а при некоторой минимальной крутизне ответы на раздражение исчезают.Это я вление принято обозначать термином « аккомодация »
В основе аккомодации лежат инактивация натриевой и повышение калиевой проводимостей,развивающиеся во время медленно нарастающей деполяризации мембраны.Аккомодация различных нервных волокон варьирует в широких пределах, но у двигательных неравных волокон скорость аккомодации,как правило,значительно выше,чем у чувствительных волокон.
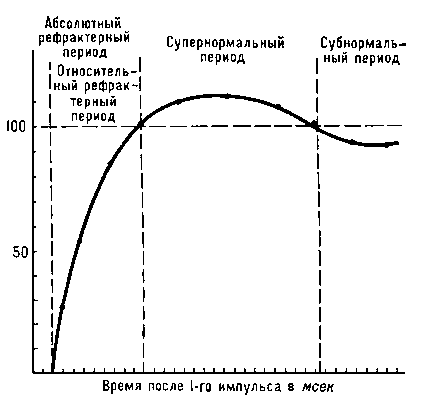
Кратковременное снижение возбудимости (См. Возбудимость) нервной и мышечной тканей непосредственно вслед за потенциалом действия (См. Потенциал действия). Р. обнаруживается при стимуляции нервов и мышц парными электрическими импульсами. Если сила 1-го импульса достаточна для возникновения потенциала действия, ответ на 2-й будет зависеть от длительности паузы между импульсами. При очень коротком интервале ответ на 2-й импульс отсутствует, как бы ни увеличивалась интенсивность стимуляции (абсолютный Рефрактерный период). Удлинение интервала приводит к тому, что 2-й импульс начинает вызывать ответ, но меньший по амплитуде, чем 1-й импульс (в опытах на нервных стволах, состоящих из большого числа параллельных нервных проводников), либо для возникновения ответа на 2-й импульс необходимо увеличить силу раздражающего тока (в опытах на одиночных нервных волокнах). Период сниженной возбудимости нервной или мышечной клетки называется относительным рефракторным периодом. За ним следует супернормальный период, или фаза экзальтации (См. Экзальтация), т. е. фаза повышенной возбудимости, сменяющаяся периодом несколько сниженной возбудимости — субнормальным периодом. В основе наблюдаемых колебаний возбудимости лежит изменение проницаемости биологических мембран (См. Проницаемость биологических мембран), сопровождающее возникновение потенциала действия (см. Биоэлектрические потенциалы). Длительность каждого периода определяется кинетикой этих процессов в данной ткани. В быстропроводящих нервных волокнах Р. длится не более 3—5 мсек, в мышце сердца период изменений возбудимости занимает до 500 мсек. Р. — один из факторов, ограничивающих частоту воспроизведения биологических сигналов, их суммацию и скорость проведения. При изменении температуры или действии некоторых лекарственных веществ длительность рефракторных периодов может меняться, чем пользуются для управления возбудимостью ткани, например сердечной мышцы: удлинение относительного рефрактерного периода приводит к снижению частоты сердечных сокращений и устранению нарушений ритма работы сердца.
Биоэлектрические явления в возбудимых тканях. Природа возбуждения
Биоэлектрические явления в возбудимых тканях. Природа возбуждения
Возбуждение представляет собой сложную совокупность физических, химических и физико-химических процессов, в результате которых происходит быстрое и кратковременное изменение электрического потенциала мембраны.
Первые исследования электрической активности живых тканей были проведены Л. Гальвани. Он обратил внимание на ссн сращение мышц препарата задних лапок лягушки, подвешенной на медном крючке, при соприкосновении с железными перилами балкона (первый опыт Гальвани). На основании этих наблюдений им был сделал вывод, что сокращение лапок вызвано «животным электричеством», которое возникает в спинном мозге и передается по металлическим проводникам (крючку и перилам) к мышцам.
Физик А. Вольта, повторив этот опыт, пришел к другому заключению. Источником тока, по его мнению, является не спинной мозг и «животное электричество», а разность потенциалов, образующаяся в месте контакта разнородных металлов – меди и железа, а нервно-мышечный препарат лягушки является лишь проводником электричества. В ответ на эти возражения Л. Гальвани усовершенствовал опыт, исключив из него металлы. Он препарировал седалищный нерв вдоль бедра лапки лягушки, затем набрасывал нерв на мышцы голени, что вызывало сокращение мышцы (второй опыт Гальвани), тем самым доказав существование «животного электричества».
Позднее Дюбуа-Реймоном было установлено, что поврежденный участок мышцы имеет отрицательный заряд, а неповрежденный участок – положительный. При набрасывании нерва между поврежденным и неповрежденным участками мышцы возникает ток, который раздражает нерв и вызывает сокращение мышцы. Этот ток был назван током покоя, или током повреждения. Так было показано, что наружная поверхность мышечных клеток заряжена положительно по отношению к внутреннему содержимому.
Мембранный потенциал
В состоянии покоя между наружной и внутренней поверхностями мембраны клетки существует разность потенциалов, которая называется мембранным потенциалом [МП), или, если это клетка возбудимой ткани, – потенциалом покоя. Так как внутренняя сторона мембраны заряжена отрицательно по отношению к наружной, то, принимая потенциал наружного раствора за нуль, МП записывают со знаком «минус». Его величина у разных клеток колеблется от минус 30 до минус 100 мВ.
Первая теория возникновения и поддержания мембранного потенциала была разработана Ю. Бернштейном (1902). Исходя из того, что мембрана клеток обладает высокой проницаемостью для ионов калия и малой проницаемостью для других ионов, он показал, что величину мембранного потенциала можно определить, используя формулу Нернста.

В 1949–1952 гг. А. Ходжкин, Э. Хаксли, Б. Катц создали со-временную мембранно-ионную теорию, согласно которой мембранный потенциал обусловлен не только концентрацией ионов калия, но и натрия и хлора, а также неодинаковой проницаемостью для этих ионов мембраны клетки. Цитоплазма нервных и мышечных клеток содержит в 30 -50 раз больше ионов калия, в 8–10 раз меньше ионов натрия и в 50 раз меньше ионов хлора, чем внеклеточная жидкость. Проницаемость мембраны для ионов обусловлена ионными каналами, макромолекулами белка, пронизывающими липидный слой. Одни каналы открыты постоянно, другие (потенциалозависимые) открываются и закрываются в ответ на изменения МП. Потенциалозависимые каналы подразделяются на натриевые, калиевые, кальциевые и хлорные. В состоянии физиологического покоя мембрана нервных клеток в 25 раз более проницаема для ионов калия, чем для ионов натрия.
Таким образом, согласно обновленной мембранной теории асимметричное распределение ионов по обе стороны мембраны и связанное с этим создание и поддержание мембранного потенциала обусловлено как избирательной проницаемостью мембраны для различных ионов, так и их концентрацией по обе стороны от мембраны, а более точно величину мембранного потенциала можно рассчитать по формуле.

Поляризация мембраны в покое объясняется наличием открытых калиевых каналов и трансмембранным градиентом концентраций калия, что приводит к выходу части внутриклеточного калия в окружающую клетку среду, т. е. к появлению положительного заряда на наружной поверхности мембраны. Органические анионы – крупномолекулярные соединения, для которых мембрана клетки непроницаема, создают на внутренней поверхности мембраны отрицательный заряд. Поэтому чем больше разница концентраций калия по обе стороны от мембраны, тем больше его выходит и тем выше значения МП. Переход ионов калия и натрия через мембрану по их концентрационному градиенту в конечном итоге должен был бы привести к выравниванию концентрации этих ионов внутри клетки и в окружающей ее среде. Но в живых клетках этого не происходит, так как в клеточной мембране имеются натрий-калиевые насосы, которые обеспечивают выведение из клетки ионов натрия и введение в нее ионов калия, работая с затратой энергии. Они принимают и прямое участие в создании МП, так как за единицу времени ионов натрия выводится из клетки больше, чем вводится калия (в соотношении 3:2), что обеспечивает постоянный ток положительных ионов из клетки. То что выведение натрия зависит от наличия метаболической энергии, доказывается тем, что под действием динитрофенола, который блокирует метаболические процессы, выход натрия снижается примерно в 100 раз. Таким образом, возникновение и поддержание мембранного потенциала обусловлено избирательной проницаемостью мембраны клетки и работой натрий-калиевого насоса.
Поделитесь на страничкеСледующая глава >
Глава 2 физиология возбудимых тканей
Способность адаптироваться к постоянно изменяющимся условиям внешней среды является одним из основных признаков живых систем. В основе приспособительных реакций организма лежит раздражимость — способность реагировать на действие различных факторов изменением структуры и функций. Раздражимостью обладают все ткани животных и растительных организмов. В процессе эволюции происходила постепенная дифференциация тканей, участвующих в приспособительной деятельности организма. Раздражимость этих тканей достигла наивысшего развития и трансформировалась в новое свойство — возбудимость. Под этим термином понимают способность ряда тканей (нервной, мышечной, железистой) отвечать на раздражение генерацией процесса возбуждения. Возбуждение — это сложный физиологический процесс временной деполяризации мембраны клеток, который проявляется специализированной реакцией ткани (проведение нервного импульса, сокращение мышцы, отделение секрета железой и т. д.). Возбудимостью обладают нервная, мышечная и секреторная ткани, которые называют возбудимыми тканями. Возбудимость различных тканей неодинакова. Ее величину оценивают по порогу раздражения — минимальной силе раздражителя, которая способна вызвать возбуждение. Менее сильные раздражители называются подпороговыми, а более сильные — сверхпороговыми.
Раздражителями, вызывающими возбуждение, могут быть любые внешние (действующие из окружающей среды) или внутренние (возникающие в самом организме) воздействия. Все раздражители по их природе можно разделить на три группы: физические (механические, электрические, температурные, звуковые, световые), химические (щелочи, кислоты и другие химические вещества, в том числе и лекарственные) и биологические (вирусы, бактерии, насекомые и другие живые существа).
По степени приспособленности биологических структур к их восприятию раздражители можно разделить на адекватные и неадекватные. Адекватными называются раздражители, к восприятию которых биологическая структура специально приспособлена в процессе эволюции. Например, адекватным раздражителем для фоторецепторов является свет, для барорецепторов — изменение давления, для мышц — нервный импульс. Неадекватными называются такие раздражители, которые действуют на структуру, специально не приспособленную для их восприятия. Например, мышца может сокращаться под влиянием механического, теплового, электрического раздражений, хотя адекватным раздражителем для нее является нервный импульс. Пороговая сила неадекватных раздражителей во много раз превышает пороговую силу адекватных.
Биоэлектрические явления в возбудимых тканях. Природа возбуждения
Возбуждение представляет собой сложную совокупность физических, химических и физико-химических процессов, в результате которых происходит быстрое и кратковременное изменение электрического потенциала мембраны.
Первые исследования электрической активности живых тканей были проведены Л. Гальвани. Он обратил внимание на сокращение мышц препарата задних лапок лягушки, подвешенной на медном крючке, при соприкосновении с железными перилами балкона (первый опыт Гальвани). На основании этих наблюдений им был сделал вывод, что сокращение лапок вызвано «животным электричеством», которое возникаете спинном мозге и передается по металлическим проводникам (крючку и перилам) к мышцам.
Физик А. Вольта, повторив этот опыт, пришел к другому заключению. Источником тока, по его мнению, является не спинной мозг и «животное электричество», а разность потенциалов, образующаяся в месте контакта разнородных металлов — меди и железа, а нервно-мышечный препарат лягушки является лишь проводником электричества. В ответ на эти возражения Л. Гальвани усовершенствовал опыт, исключив из него металлы. Он препарировал седалищный нерв вдоль бедра лапки лягушки, затем набрасывал нерв на мышцы голени, что вызывало сокращение мышцы (второй опыт Гальвани), тем самым доказав существование «животного электричества».
Позднее Дюбуа-Реймоном было установлено, что поврежденный участок мышцы имеет отрицательный заряд, а неповрежденный участок — положительный. При набрасывании нерва между поврежденным и неповрежденным участками мышцы возникает ток, который раздражает нерв и вызывает сокращение мышцы. Этот ток был назван током покоя, или током повреждения. Так было показано, что наружная поверхность мышечных клеток заряжена положительно по отношению к внутреннему содержимому.
Лекция . Биоэлектричество и его механизмы.
8
План лекции.
История открытия биоэлектричества. Токи покоя и действия.
Мембранный потенциал покоя. Методы регистрации.
Природа мембранного потенциала покоя.
Изменение мембранного потенциала при возбуждении (потенциал действия (ПД)).
Сравнительная характеристика ЛО и ПД.
Современные представления о механизме ПД.
Потенциал действия и возбудимости клетки.
1. Первые научные работы, направленные на изучение «животного электричества», датированы серединой 18 столетия и связаны с изучением электрических органов морских животных (электрический скат, некоторые глубоководные рыбы). Все эти исследования подготовили благоприятную почву для работ Л.Гальвани, который считается основателем электрофизиологии. Само открытие «животного электричества» у лягушек связано с так называемым «балконным опытом» (1786). Должен отметить, что на первом этапе своей научной карьеры Л.Гальвани изучал влияние различных по природе электрических разрядов на нервно-мышечный препарат и препарат задних лапок лягушек. При подготовке эксперимента он обнаружил любопытное явление: касание препарата задних лапок лягушки, висящего на медном крючке, железных прутьев балкона, сопровождалось сокращением мышц. Исследователь предположил, что между мышечной и нервной тканью препарата существует разность потенциалов: сокращение мышц происходит в результате замыкания электрической цепи, где медный крючок и железный прут выступают в роли проводников. Современник Л.Гальвани известный физик А.Вольта трактовал это явление как результат возникновения постоянного тока в цепи, состоящей из разнородных металлов. В роли проводника, по его мнению, выступал препарат. Сегодня мы знаем, что в данном конкретном случае был прав А.Вольта, однако, дальнейшими экспериментами Л.Гальвани и его ученики доказали, что «животное электричество» существует (второй опыт Л.Гальвани с исключением контакта разнородных металлов, опыт К.Маттеуччи с двумя нервно-мышечными препаратами, один из которых при возбуждении электрическим током служил источником возбуждения для другого).
Второй исторический этап развития электрофизиологии связывают с именем выдающегося немецкого физиолога Эмиля Дю Буа-Реймона. В 1841 он показал, что при замыкании измерительной электрической цепи между поврежденным и неповрежденным участком мышцы, можно зарегистрировать электрический ток, названный им «током покоя». При возбуждении мышцы величина величина «тока покоя» уменьшалась. Это колебание «тока покоя» он назвал «током действия». Опыт Э.Дю Буа-Реймона позволял предположить, что между поверхностью мышечной клетки и ее протоплазмой в состоянии покоя существует разность потенциалов. Действительно, при пересечении мышцы мы обеспечиваем в эксперименте доступ к протоплазме мышечных клеток, а помещая в этот участок электрод и соединяя его с гальванометром, второй вход которого соединен с неповрежденным участком, экспериментатор оценивает разность потенциалов между указанными точками. Развитие электрофизиологии в 40-50 годы нашего столетия подтвердило это предположение.
2. Современная материально-техническая электрофизиологическая база позволяет осуществить регистрацию разности потенциалов в покоящейся возбудимой клетке. Для этого необходимы: микроэлектроды с сечением 0,5 мкм (металлические или стеклянные), усилитель постоянного тока и осциллограф (см.схему 1). Один из микроэлектродов вводится внутрь клетки, а другой помещается на ее поверхность. Электроды соединяются со входами усилителя, выходы которого подаются на вход осциллографа (рис.1).

УСИЛИТЕЛЬ ПОСТОЯННОГО ТОКА





ОСЦИЛЛОГРАФ
ВОЗБУДИМАЯ
КЛЕТКА
Рис.1. Схема экспериментальной установки для регистрации МПП.
На экране осциллографа разность потенциалов между поверхностью мембраны клетки и ее протоплазмой отражается смещением луча от изолинии, причем чем больше это отклонение, тем больше величина разности потенциалов.
3. Первую гипотезу в отношении природы МПП высказал Э.Дю Буа-Реймон, предположивший, что разность потенциалов создается молекулами, встроенными в мембрану клеток и имеющими электростатические заряды на полюсах (дипольная гипотеза). Однако, эта гипотеза не была подтверждена экспериментально и сегодня представляет лишь исторический интерес.
Дальнейшее развитие представлений в области биоэлектричества связано с именем В.Ю.Чаговца, высказавшем гипотезу об ионной природе этих явлений (1896). В частности, он считал, что в процессе жизнедеятельности клеток синтезируются различные органические кислоты, которые согласно теории электролитической диссоциации Аррениуса в водной фазе протоплазмы клеток диссоциируют на ионы водорода и анионы. Положительно заряженные ионы водорода более подвижны, чем анионы, в связи с чем они выходят на поверхность клеток, формируя положительный заряд на них. Анионы же остаются в протоплазме и формируют ее электроотрицательность. Однако, эта гипотеза не нашла своего подтверждения: при таком механизме формированияразности потенциалов в клетках рН межклеточной жидкости должна быть кислой, тогда как в реальности она нейтральная. Кроме того абсолютное значение разности потенциалов, согласно этой гипотезе, должно быть меньшей, чем установлено экспериментально. На следующем этапе изучения электрогенеза в тканях возникла классическая мембранно-ионная теория, автором которой был один из учеников Э.Дю Буа-Реймона Ю.Берштейн (1902). Согласно его взглядам, ведущими факторами формирования разности потенциалов в клетке в состоянии покоя является ее полупроницаемая мембрана и ионная асимметрия. В своих представлениях Ю.Берштейн опирался на опыт Оствальда, демонстрирующего появление разности потенциалов между двумя половинами сосуда с раствором К2SО, перегороженного полупроницаемой мембраной. Величина этой разности потенциалов должна соответствовать расчетной на основе формулы Нернста:
Е= ((R*Т) / ( F*n )) * Ln (К+ Н / К+ В),
где Е- разность потенциалов между наружной и внутренней поверхностью мембраны, R- газовая постоянная, Т – абсолютная температура,F- число Фарадея ,К+ Н и К+ В –внутренняя и наружная концентрация ионов калия (число Фарадея – заряд 1 моля одновалентных ионов, газовая постоянная – кинетическая энергия одного моля ионов при Т=1К). В целом верно отражая механизм электрогенеза, теория Ю.Берштейна не во всех случаях корректно объясняла электрофизиологические феномены, например, происхождение части высоковольтного пика – овершута, не объясняла причину возникновения исходных ионных градиентов.
Развитие представлений Ю.Берштейна нашло в работах английских физиологов Эндрю Ф.Хаксли, Алана Л.Ходжкина и Бернарда Катца. Согласно их взглядам, природу МПП можно объяснить, базируясь на следующих положениях.
Лекция 1 Введение в физиологию Физиология возбудимых тканей
 Лекция 1 Введение в физиологию. Физиология возбудимых тканей
Лекция 1 Введение в физиологию. Физиология возбудимых тканей
 Физиология – наука, изучающая функционирование живых организмов, их отдельных систем, органов, тканей и клеток, а также их взаимодействие с окружающей средой
Физиология – наука, изучающая функционирование живых организмов, их отдельных систем, органов, тканей и клеток, а также их взаимодействие с окружающей средой
 • Физиологическая функция – специфическое проявление жизнедеятельности организма или его частей и имеющее приспособительное значение. • Физиологическая система – термин, описывающий условную живую биологическую систему на любом уровне организации живой материи, начиная с клеточного уровня и включая его.
• Физиологическая функция – специфическое проявление жизнедеятельности организма или его частей и имеющее приспособительное значение. • Физиологическая система – термин, описывающий условную живую биологическую систему на любом уровне организации живой материи, начиная с клеточного уровня и включая его.
 Основные разделы современной физиологии • общая • частная -физиология дыхания -физиология кровообращения и т. д. • прикладная -физиология труда -физиология спорта -физиология питания -экологическая физиология
Основные разделы современной физиологии • общая • частная -физиология дыхания -физиология кровообращения и т. д. • прикладная -физиология труда -физиология спорта -физиология питания -экологическая физиология
 • В общую физиологию включают сведения, • касающиеся природы основных жизненных процессов, общих проявлений жизнедеятельности, таких как метаболизм органов и тканей, свойства биологических мембран и отдельных клеток, общие закономерности реагирования организма и его структур на воздействие среды — раздражимость, возбудимость, процессы возбуждения и торможения. Сюда же относят особенности жизнедеятельности, обусловленные уровнем структурной организации, разными условиями существования и целым рядом других причин. Частная физиология исследует свойства отдельных клеток, тканей (мышечной, нервной и др. ), органов (печени, почек, сердца и пр. ), закономерности объединения их в системы (система пищеварения, кровообращения, дыхания), а также физиологию отдельных классов, групп и видов животных. Прикладная физиология изучает закономерности проявлении деятельности организма, особенно человека, в связи со специальными задачами и условиями.
• В общую физиологию включают сведения, • касающиеся природы основных жизненных процессов, общих проявлений жизнедеятельности, таких как метаболизм органов и тканей, свойства биологических мембран и отдельных клеток, общие закономерности реагирования организма и его структур на воздействие среды — раздражимость, возбудимость, процессы возбуждения и торможения. Сюда же относят особенности жизнедеятельности, обусловленные уровнем структурной организации, разными условиями существования и целым рядом других причин. Частная физиология исследует свойства отдельных клеток, тканей (мышечной, нервной и др. ), органов (печени, почек, сердца и пр. ), закономерности объединения их в системы (система пищеварения, кровообращения, дыхания), а также физиологию отдельных классов, групп и видов животных. Прикладная физиология изучает закономерности проявлении деятельности организма, особенно человека, в связи со специальными задачами и условиями.
 Прикладная физиология • К числу наиболее актуальных для медицины относят клиническую физиологию, изучающую причины возникновения • отклонений физиологических функций от их нормативных значений. Также здесь следует упомянуть физиологию труда, физиология спорта, физиологии питания. Ряд специальных проблем, возникающих в результате пребывания организма при измененном барометрическом давлении, перегрузках, условиях, не встречающихся на Земле (невесомость), изучает физиология подводного плавания, авиационная и космическая физиология. • Влияния, оказываемые на организм внешней средой, и связанные • • с ними особенности физиологических процессов у разных видов животных в зависимости от условий существования изучает по преимуществу экологическая физиология. сравнительная физиология изучает функции отдельных органов и систем у самых разных организмов, выявляя общие принципы и закономерности их функциональной организации. Кроме того, существуют такие разделы как эволюционная физиология, физиология адаптации.
Прикладная физиология • К числу наиболее актуальных для медицины относят клиническую физиологию, изучающую причины возникновения • отклонений физиологических функций от их нормативных значений. Также здесь следует упомянуть физиологию труда, физиология спорта, физиологии питания. Ряд специальных проблем, возникающих в результате пребывания организма при измененном барометрическом давлении, перегрузках, условиях, не встречающихся на Земле (невесомость), изучает физиология подводного плавания, авиационная и космическая физиология. • Влияния, оказываемые на организм внешней средой, и связанные • • с ними особенности физиологических процессов у разных видов животных в зависимости от условий существования изучает по преимуществу экологическая физиология. сравнительная физиология изучает функции отдельных органов и систем у самых разных организмов, выявляя общие принципы и закономерности их функциональной организации. Кроме того, существуют такие разделы как эволюционная физиология, физиология адаптации.
 • Нормальная физиология — является фундаментальной базой практически всех медицинских и ветеринарных дисциплин. Несомненно, без знания нормального течения патологических процессов нельзя лечить больного. • Патологическая физиология (патофизиология) рассматривает измененные функции больного организма, выясняет общие закономерности возникновения, развития и течения патологических процессов в организме, процессы компенсации и адаптации функций при заболеваниях, а также механизмы выздоровления и реабилитации.
• Нормальная физиология — является фундаментальной базой практически всех медицинских и ветеринарных дисциплин. Несомненно, без знания нормального течения патологических процессов нельзя лечить больного. • Патологическая физиология (патофизиология) рассматривает измененные функции больного организма, выясняет общие закономерности возникновения, развития и течения патологических процессов в организме, процессы компенсации и адаптации функций при заболеваниях, а также механизмы выздоровления и реабилитации.
 • Физиология – это сугубо экспериментальная наука, основным методом познания механизмов и закономерностей в ней является эксперимент, позволяющий не только ответить на вопрос, что происходит в организме, но и выяснить также, как и почему происходит тот или иной физиологический процесс, как он возникает, какими механизмами поддерживается и управляется.
• Физиология – это сугубо экспериментальная наука, основным методом познания механизмов и закономерностей в ней является эксперимент, позволяющий не только ответить на вопрос, что происходит в организме, но и выяснить также, как и почему происходит тот или иной физиологический процесс, как он возникает, какими механизмами поддерживается и управляется.
 Методы исследования • Острые • Хронические — Аналитические — Синтетические • • • Экстирпации Трансплантации Денервации Стимуляции Фистульный Пересадки органа Клинико-анатомические Инструментальные Генетические
Методы исследования • Острые • Хронические — Аналитические — Синтетические • • • Экстирпации Трансплантации Денервации Стимуляции Фистульный Пересадки органа Клинико-анатомические Инструментальные Генетические
 • Острый эксперимент обычно • непродолжителен. В этом случае наркотизированное и обездвиженное животное вскрывают для проведения искусственной изоляции органов и тканей, иссечения и стимуляции различных нервов, регистрации электрических потенциалов, введения лекарственных препаратов и т. д. Хронический эксперимент требует специальной подготовки в виде определенно направленных хирургических операций и использования животного в опыте только после того, как оно оправится от хирургического вмешательства.
• Острый эксперимент обычно • непродолжителен. В этом случае наркотизированное и обездвиженное животное вскрывают для проведения искусственной изоляции органов и тканей, иссечения и стимуляции различных нервов, регистрации электрических потенциалов, введения лекарственных препаратов и т. д. Хронический эксперимент требует специальной подготовки в виде определенно направленных хирургических операций и использования животного в опыте только после того, как оно оправится от хирургического вмешательства.
 инвазивные и неинвазивные методы • . Инвазивные – методы исследования функций физиологической системы, которые сопровождаются ее частичным повреждением (например, взятие пробы крови или ликвора), • неинвазивными именуют методики при которых, не происходит повреждения или нарушения целостности физиологической системы (например, электрокардиография или электроэнцефалография).
инвазивные и неинвазивные методы • . Инвазивные – методы исследования функций физиологической системы, которые сопровождаются ее частичным повреждением (например, взятие пробы крови или ликвора), • неинвазивными именуют методики при которых, не происходит повреждения или нарушения целостности физиологической системы (например, электрокардиография или электроэнцефалография).
 инструментальные методы • Электрофизиологические • Клинические и функциональные пробы • Functional brain imaging -Позитронно-эмиссионная томография -Однофотонная эмиссионная компьютерная томография -functional magnetic resonance imaging
инструментальные методы • Электрофизиологические • Клинические и функциональные пробы • Functional brain imaging -Позитронно-эмиссионная томография -Однофотонная эмиссионная компьютерная томография -functional magnetic resonance imaging
 functional magnetic resonance imaging
functional magnetic resonance imaging
") Метод нокаута генов • позволяет получать линии нокаутных мышей (knock- out mice, knockout mice) — мутантных мышей, у которых выключены определенные гены. Этот метод позволяет исследовать роль каждого конкретного гена в развитии организма и в его нормальной и патологической работе и изучать различные человеческие болезни, используя мышей в качестве модельных объектов. Выключенный ген приводит к тем или иным нарушениям. Характер этих нарушений позволяет судить о функциях данного гена.
Метод нокаута генов • позволяет получать линии нокаутных мышей (knock- out mice, knockout mice) — мутантных мышей, у которых выключены определенные гены. Этот метод позволяет исследовать роль каждого конкретного гена в развитии организма и в его нормальной и патологической работе и изучать различные человеческие болезни, используя мышей в качестве модельных объектов. Выключенный ген приводит к тем или иным нарушениям. Характер этих нарушений позволяет судить о функциях данного гена.
 Методы • клинические или исследования человека • (применяются в клинической работе врача и достаточно ограничены по своим возможностям). Например, электрокардиография или электроэнцефалография — экспериментальные или эксперименты на животных (применяются в эксперименте на животных). Чрезвычайно многообразны и лежат в основе возникновения новых клинических методов. В свою очередь экспериментальные методы можно классифицировать по форме проведения физиологического эксперимента на «острые» и хронические.
Методы • клинические или исследования человека • (применяются в клинической работе врача и достаточно ограничены по своим возможностям). Например, электрокардиография или электроэнцефалография — экспериментальные или эксперименты на животных (применяются в эксперименте на животных). Чрезвычайно многообразны и лежат в основе возникновения новых клинических методов. В свою очередь экспериментальные методы можно классифицировать по форме проведения физиологического эксперимента на «острые» и хронические.
 - это относительно") Основные свойства живого организма • живой организм (или физиологическая • система) — это относительно автономная, самостоятельно функционирующая, саморегулирующаяся система, реагирующая как единое целое на изменение среды основные свойства живых систем метаболизм, рост и развитие, способность к размножению, целостность и дискретность, раздражимость и возбудимость, гомеостаз и др.
Основные свойства живого организма • живой организм (или физиологическая • система) — это относительно автономная, самостоятельно функционирующая, саморегулирующаяся система, реагирующая как единое целое на изменение среды основные свойства живых систем метаболизм, рост и развитие, способность к размножению, целостность и дискретность, раздражимость и возбудимость, гомеостаз и др.
 Период грудного") ВОЗРАСТНЫЕ ПЕРИОДЫ РАЗВИТИЯ РЕБЕНКА • • • Период новорожденности (3— 4 недели) Период грудного возраста (до 1 года) Ранний детский возраст (от 1 года до 3 -х лет) Дошкольный возраст (от 3 до 7 лет) Младший школьный возраст (от 7 до 12 лет) Старший школьный возраст (у девочек с 11 — 13 до 16— 18 лет, у мальчиков с 15— 16 лет до 19— 20 лет)
ВОЗРАСТНЫЕ ПЕРИОДЫ РАЗВИТИЯ РЕБЕНКА • • • Период новорожденности (3— 4 недели) Период грудного возраста (до 1 года) Ранний детский возраст (от 1 года до 3 -х лет) Дошкольный возраст (от 3 до 7 лет) Младший школьный возраст (от 7 до 12 лет) Старший школьный возраст (у девочек с 11 — 13 до 16— 18 лет, у мальчиков с 15— 16 лет до 19— 20 лет)
 механизмы регуляции функций организма • Нервный. Характеризуется меньшим • латентным периодом, большей скоростью, локальностью ответа. Гуморальный – за счет циркуляции в крови определенных химических соединений. Характеризуется большим латентным периодом, меньшей скоростью, и диффузностью ответа.
механизмы регуляции функций организма • Нервный. Характеризуется меньшим • латентным периодом, большей скоростью, локальностью ответа. Гуморальный – за счет циркуляции в крови определенных химических соединений. Характеризуется большим латентным периодом, меньшей скоростью, и диффузностью ответа.
 всех • физиологических") Базовые понятия физиологии возбудимых систем • Раздражимость – это способность (свойство) всех • физиологических систем активно отвечать на внешнее или внутреннее воздействие той или иной ответной реакцией, например, усилением метаболизма и роста, ускорением деления, выбросом секрета, движением и т. д. Возбудимость – это способность (свойство) некоторых физиологических систем отвечать на внешнее или внутреннее воздействие в виде специализированной ответной реакции – генерации потенциала действия.
Базовые понятия физиологии возбудимых систем • Раздражимость – это способность (свойство) всех • физиологических систем активно отвечать на внешнее или внутреннее воздействие той или иной ответной реакцией, например, усилением метаболизма и роста, ускорением деления, выбросом секрета, движением и т. д. Возбудимость – это способность (свойство) некоторых физиологических систем отвечать на внешнее или внутреннее воздействие в виде специализированной ответной реакции – генерации потенциала действия.
 раздражители • Пороговые • Подпороговые • Надпороговые • Физические • Химические • Биологические • Адекватные • Неадекватные
раздражители • Пороговые • Подпороговые • Надпороговые • Физические • Химические • Биологические • Адекватные • Неадекватные
 Кривая «сила-длительность» • АВ – реобаза • АЕ – полезное • • время АД – хронаксия АС – удвоенная реобаза
Кривая «сила-длительность» • АВ – реобаза • АЕ – полезное • • время АД – хронаксия АС – удвоенная реобаза
 Строение мембран
Строение мембран
 Липиды мембран • Холестерол • Гликолипиды -ганглиозиды -цереброзиды • Фосфолипиды -сфингомиелин -фосфатидилхолин -фосфатидилсерин -этаноламин
Липиды мембран • Холестерол • Гликолипиды -ганглиозиды -цереброзиды • Фосфолипиды -сфингомиелин -фосфатидилхолин -фосфатидилсерин -этаноламин
 функции мембран 1. Барьерная 2. Метаболическая 3. Электрогенная 4. Иммуннологическая 5. Рецептивная 6. Обеспечение межклеточного взаимодействия.
функции мембран 1. Барьерная 2. Метаболическая 3. Электрогенная 4. Иммуннологическая 5. Рецептивная 6. Обеспечение межклеточного взаимодействия.
 Транспорт веществ через мембрану • Пассивный • -Простая диффузия -Облегченная диффузия -Осмос Активный -Первично-активный -Вторично-активный а) Симпорт б) Антипорт
Транспорт веществ через мембрану • Пассивный • -Простая диффузия -Облегченная диффузия -Осмос Активный -Первично-активный -Вторично-активный а) Симпорт б) Антипорт

 Классификация ионных каналов • По типу активации -Потенциалзависимые -Лигандзависимые • По селективности -Селективные (Na, K, Ca, Cl) -Неселективные • По кинетике -Быстрые -Медленные
Классификация ионных каналов • По типу активации -Потенциалзависимые -Лигандзависимые • По селективности -Селективные (Na, K, Ca, Cl) -Неселективные • По кинетике -Быстрые -Медленные
 Вторично-активный транспорт
Вторично-активный транспорт


 Потенциалзависимые каналы
Потенциалзависимые каналы
 Лигандзависимые каналы
Лигандзависимые каналы

 Молекулярная структура ионных каналов
Молекулярная структура ионных каналов
 Схема работы воротного механизма Na канала
Схема работы воротного механизма Na канала

 Мембранный потенциал б) Потенциал") Электрические процессы в возбудимых тканях • 1. Токи покоя а) Мембранный потенциал б) Потенциал повреждения в) Метаболический ток • 2. Токи действия
Электрические процессы в возбудимых тканях • 1. Токи покоя а) Мембранный потенциал б) Потенциал повреждения в) Метаболический ток • 2. Токи действия
 Внутри Снаружи Катионы ы+") Распределение ионов снаружи и внутри клетки Ионы Концентрация (м. М/л) Внутри Снаружи Катионы ы+ 124 10 10 5 0, 005 2 2 125 2 2 Анионы 2 12 139 77 27 26 K+ K + Na 2+ Ca Cl- HCO 3 Орг. анион ы
Распределение ионов снаружи и внутри клетки Ионы Концентрация (м. М/л) Внутри Снаружи Катионы ы+ 124 10 10 5 0, 005 2 2 125 2 2 Анионы 2 12 139 77 27 26 K+ K + Na 2+ Ca Cl- HCO 3 Орг. анион ы
 Основные причины формирования мембранного потенциала • Избирательная проницаемость мембраны • Ионная асимметрия
Основные причины формирования мембранного потенциала • Избирательная проницаемость мембраны • Ионная асимметрия
 Уравнение Гольдмана
Уравнение Гольдмана
 Уравнение Нернста
Уравнение Нернста
 Эквивалентная электрическая схема мембраны
Эквивалентная электрическая схема мембраны

 Потенциал действия
Потенциал действия
 Фазы потенциала действия • Деполяризации • Реполяризации • Отрицательный следовой потенциал • Положительный следовой потенциал
Фазы потенциала действия • Деполяризации • Реполяризации • Отрицательный следовой потенциал • Положительный следовой потенциал
 Na-K насос
Na-K насос
 ИЗМЕНЕНИЯ ВОЗБУДИМОСТИ ПРИ ВОЗБУЖДЕНИИ Фазы возбуждения: 1. Абсолютной рефрактерности 2. Относительной рефрактерности 3. Повышенной возбудимости 4. Сниженной возбудимости
ИЗМЕНЕНИЯ ВОЗБУДИМОСТИ ПРИ ВОЗБУЖДЕНИИ Фазы возбуждения: 1. Абсолютной рефрактерности 2. Относительной рефрактерности 3. Повышенной возбудимости 4. Сниженной возбудимости