


МИГРАЦИЯ СЕЛЕКТИВНАЯ — это… Что такое МИГРАЦИЯ СЕЛЕКТИВНАЯ?
- МИГРАЦИЯ СЕЛЕКТИВНАЯ
-
— англ. migration, selective; нем. Wanderung, selektive. Миграция представителей определенной категории или группы лиц того или иного соц. класса или слоя, территориальных и этнических групп, нац. меньшинств и др.
Antinazi. Энциклопедия социологии, 2009
- МИГРАЦИЯ СЕЗОННАЯ
- МИД
Смотреть что такое «МИГРАЦИЯ СЕЛЕКТИВНАЯ» в других словарях:
Функции миграции населения — это те конкретные роли, которые играют миграции населения в жизнедеятельности общества. Естественно, что функции миграции выражают ее сущность, свойства этого явления.
 Функции миграции населения независимы от типа социально экономической системы… … Миграция: словарь основных терминов
Функции миграции населения независимы от типа социально экономической системы… … Миграция: словарь основных терминовФУНКЦИИ МИГРАЦИИ — роли выполняемые миграцией в об ве. К общим функциям миграции относятся: развитие подвижности, перераспределительная и селективная функция. Первая функция состоит в обеспечении того или иного уровня пространственной подвижности, к рая… … Российская социологическая энциклопедия
Экономика Белоруссии — Экономика Республики Беларусь Экономические показатели Валюта Белорусский рубль (BYR) Фискальный год календарный Международные организации СНГ, ЕврАзЭС Статистика ВВП (номинальный) $54,7 млрд. (2010, оценка … Википедия
«ЧЕЛОВЕЧЕСКОГО КАПИТАЛА» ТЕОРИЯ
ИЗОТОПОВ РАЗДЕЛЕНИЕ, — ИЗОТОПОВ РАЗДЕЛЕНИЕ, выделение отдельных изотопов из естеств.
их смеси или обогащение смеси отдельными изотопами. Первые попытки И. р. сделаны Ф. У. Астоном (F. W. Aston, 1949) и др. гл. обр. для обнаружения изотопов у стабильных элементов,… … Физическая энциклопедия
 Функции миграции населения независимы от типа социально экономической системы… … Миграция: словарь основных терминов
Функции миграции населения независимы от типа социально экономической системы… … Миграция: словарь основных терминов их смеси или обогащение смеси отдельными изотопами. Первые попытки И. р. сделаны Ф. У. Астоном (F. W. Aston, 1949) и др. гл. обр. для обнаружения изотопов у стабильных элементов,… … Физическая энциклопедия
их смеси или обогащение смеси отдельными изотопами. Первые попытки И. р. сделаны Ф. У. Астоном (F. W. Aston, 1949) и др. гл. обр. для обнаружения изотопов у стабильных элементов,… … Физическая энциклопедияРешение проблем селективной выборки путем декластеризации данных—ArcMap
Доступно с лицензией Geostatistical Analyst.
Часто данные расположены в пространстве и не случайным образом, и не по регулярной сетке. По разным причинам выборка данных может быть селективной, с более высокой плотностью элементов выборки в некоторых местах. Это важно учитывать для надлежащего выполнения преобразования по методу нормальных меток и для того, чтобы гистограмма (а также интегральная функция распределения) выборки правильно отражала гистограмму всей генеральной совокупности. В случае селективной выборки данных при наличии пространственной автокорреляции получившаяся гистограмма выборки может не отражать гистограмму генеральной совокупности.
В выборке декластеризации на левом верхнем рисунке генеральная совокупность значений в 100 точках вдоль линии представлена в виде закрашенных окружностей. Это результат моделирования из процесса с пространственной автокорреляцией, постоянным средним и устойчивой положительной автокорреляцией. В качестве выборки используется каждая вторая точка, начиная с первой. Точки выборки представлены в виде окружностей. Справа в выборке декластеризации синим цветом показана гистограмма генеральной совокупности и фиолетовым цветом – гистограмма выборки.
Поскольку выборка – это половина генеральной совокупности, можно ожидать, что высота столбцов гистограммы выборки будет примерно вдвое меньше, чем у гистограммы генеральной совокупности, с некоторыми отклонениями. Слева внизу приведена селективная выборка данных: выбрана каждая пятая точка до точки 34, затем каждая точка до точки 70 и снова каждая пятая точка до конца. В результате опять получена выборка, представляющая половину генеральной совокупности. В результате предпочтительной выборки в средней области пространственных местоположений в выборке будет представлен более высокий процент средних значений, поэтому высота столбцов гистограммы выборки будет практически равна высоте столбцов гистограммы генеральной совокупности для значений от -3 до 1. В то же время более низкие и более высокие значения будут недостаточно представлены в гистограмме выборки.
В результате предпочтительной выборки в средней области пространственных местоположений в выборке будет представлен более высокий процент средних значений, поэтому высота столбцов гистограммы выборки будет практически равна высоте столбцов гистограммы генеральной совокупности для значений от -3 до 1. В то же время более низкие и более высокие значения будут недостаточно представлены в гистограмме выборки.
Одно из решений проблем, связанных с селективной выборкой, – использование для данных весовых коэффициентов, когда в областях плотной выборки данным присваиваются меньшие веса (в результате в приведенном выше примере селективной выборки столбцы гистограммы выборки уменьшатся для значений от -3 до 1), а в областях редкой выборки – более высокие веса (в результате столбцы гистограммы выборки станут шире для более высоких и более низких значений). В ArcGIS Geostatistical Analyst Extension допустимы два метода. По умолчанию применяется метод декластеризации ячеек. В этом случае прямоугольные ячейки размещаются поверх местоположений данных в сетке, а весовой коэффициент, присваиваемый каждому местоположению данных, обратно пропорционален количеству точек данных в соответствующей ячейке.
Остается только выбрать размер и ориентацию сетки. В ArcGIS Geostatistical Analyst Extension предусмотрена диаграмма, которая показывает средневзвешенное значение среди всех данных для разных размеров ячеек. Рекомендуется выбирать размер ячейки в соответствии с минимальным средневзвешенным значением в случае селективной выборки данных в областях высоких значений, и наоборот, выбирать размер ячейки в соответствии с максимальным средневзвешенным значением в случае селективной выборки данных в областях низких значений.
В другой схеме используется полигональный метод, определяющий полигон вокруг каждого пространственного местоположения данных таким образом, что все точки внутри этого полигона расположены ближе к местоположению данных, чем любые другие местоположения, как показано на следующем рисунке.
Местоположения данных показаны маленькими точками, вокруг них нарисованы полигоны с цветной штриховкой, показывающей размер полигонов. Идея заключается в назначении веса каждому местоположению данных пропорционально области, которую оно «представляет».
Более подробно о декластеризации для решения проблем селективной выборки
Роданидная функция тетрароданоцинкат- селективного электрода на основе тринонилоктадециламмония
Please use this identifier to cite or link to this item:
https://elib.bsu.by/handle/123456789/13845
| Title: | Роданидная функция тетрароданоцинкат- селективного электрода на основе тринонилоктадециламмония |
| Authors: | Рахманько, Е. М. М.Матвейчук, Ю. В. Ясинецкий, В. В. |
| Keywords: | ЭБ БГУ::ЕСТЕСТВЕННЫЕ И ТОЧНЫЕ НАУКИ::Химия |
| Issue Date: | Jun-2011 |
| Publisher: | БГУ |
| Citation: | Вестник Белорусского государственного университета. Сер. 2, Химия. Биология. География. – 2011. — № 2. – С. 10-14. |
| Abstract: | The occurrence of the ligand (thiocyanate) response of the tetrathiocyanozincate-selective film electrode based on a higher
quaternary ammonium salt (TNODA+I–) was explained by the exchange replacement of zinc thiocyanate ions by thiocyanate ions
from the membrane to the near-electrode layer. The determination limit of the electrode was explained by the dissociation of the
ionophore. Ligand (thiocyanate) response of the tetrathiocyanozincate-selective film electrode on the background of high
concentrations ZnCl2 have high slope and more low limit of revelation, that is perspective for determination of thiocyanate-ions. |
| URI: | http://elib.bsu.by/handle/123456789/13845 |
| ISSN: | 0372-5340 |
| Appears in Collections: | 2011, №2 (июнь) |
Items in DSpace are protected by copyright, with all rights reserved, unless otherwise indicated.
Селективные XML-индексы — SQL Server
- Чтение занимает 5 мин
В этой статье
Применимо к: SQL Server (все поддерживаемые версии) База данных SQL Azure
Селективные XML-индексы — это еще один тип XML-индексов, доступный наряду с обычными XML-индексами. Селективный XML-индекс используется в следующих целях.
Селективный XML-индекс используется в следующих целях.
Для повышения производительности запросов к XML-данным, хранящимся в SQL Server.
Для поддержки более быстрого индексирования больших объемов XML-данных.
Для повышения масштабируемости путем уменьшения затрат на хранение XML-индексов.
Основным ограничением обычных XML-индексов является то, что они индексируют весь XML-документ. Это ограничение влечет за собой несколько значительных недостатков, например снижение производительности запросов и увеличенные затрат на обслуживание, в основном относящихся к затратам на хранение индекса.
Функция селективного XML-индекса позволяет повышать только некоторые пути из XML-документов в индекс. Во время создания индекса производится вычисление этих путей, а узлы, на которые они указывают, разделяются и сохраняются в реляционной таблице в SQL Server. Эта функция использует эффективный алгоритм сопоставления, разработанный Microsoft Research в сотрудничестве с группой разработчиков SQL Server . Этот алгоритм сопоставляет узлы XML с одной реляционной таблицей и достигает исключительной производительности при использовании небольшого пространства для хранения.
Этот алгоритм сопоставляет узлы XML с одной реляционной таблицей и достигает исключительной производительности при использовании небольшого пространства для хранения.
Функция селективного XML-индекса также поддерживает вторичные селективные XML-индексы для узлов, которые были индексированы селективным XML-индексом. Эти вторичные селективные индексы являются эффективными и еще более повышают производительность запросов.
Преимущества селективных XML-индексов
Селективные XML-индексы дают следующие преимущества:
Значительное улучшение производительности запросов к типу данных XML для типичных нагрузок по запросам.
Сниженные требования к хранилищу по сравнению с обычными XML-индексами.
Сниженные расходы на обслуживание индекса по сравнению с обычными XML-индексами.
Нет необходимости обновлять приложения для получения пользы от селективных XML-индексов.
Селективные XML-индексы и первичные XML-индексы
Важно!
Создание селективного XML-индекса вместо обычного XML-индекса в большинстве случаев приводит к повышению производительности и более эффективному использованию хранилища.
Однако селективный XML-индекс не рекомендуется использовать при наличии следующих условий.
Необходимо сопоставить большое число путей узлов.
Поддерживаются запросы неизвестных элементов или элементов в неизвестном месте структуры документа.
Простой пример селективного XML-индекса
Рассмотрим следующий фрагмент XML как XML-документ в таблице из приблизительно 500 000 строк.
<book>
<created>2004-03-01</created>
<authors>Various</authors>
<subjects>
<subject>English wit and humor -- Periodicals</subject>
<subject>AP</subject>
</subjects>
<title>Punch, or the London Charivari, Volume 156, April 2, 1919</title>
<id>etext11617</id>
</book>
Создание первичного XML-индекса для такого большого количества строк этой простой схемы занимает длительное время. Создание запросов к этим данным также является неэффективным, поскольку первичный XML-индекс не поддерживает селективное индексирование.
Создание запросов к этим данным также является неэффективным, поскольку первичный XML-индекс не поддерживает селективное индексирование.
Если запросы к данным выполняются только по путям /book/title и /book/subjects , вы можете создать следующий селективный XML-индекс.
CREATE SELECTIVE XML INDEX SXI_index
ON Tbl(xmlcol)
FOR
(
pathTitle = '/book/title/text()' AS XQUERY 'xs:string',
pathAuthors = '/book/authors' AS XQUERY 'node()',
pathId = '/book/id' AS SQL NVARCHAR(100)
)
Предыдущая инструкция — это хороший пример синтаксиса CREATE, используемого при создании селективного XML-индекса. В инструкции CREATE указывается имя индекса и определяется таблица и XML-столбец для индексирования. Затем передаются пути для индексирования. Путь состоит из 3 частей:
имя, однозначно определяющее путь;
выражение XQuery, описывающее путь;
необязательные указания по оптимизации.

Дополнительные сведения об этих элементах см. в разделе Связанные задачи.
Поддерживаемые функции, предварительные требования и ограничения
Поддерживаемые функции XML
Селективные XML-индексы поддерживают XQuery, поддерживаемые SQL Server в методах exist(), value() и nodes().
Для методов exist(), value() и nodes() селективные XML-индексы содержат достаточно данных для преобразования всего выражения.
Для методов query() и modify() селективные XML-индексы можно использовать только для фильтрации узлов.
Для метода query() селективные XML-индексы не используются для получения результатов.
Для метода modify() селективные XML-индексы не используются для обновления XML-документов.
Неподдерживаемые функции XML
Селективные XML-индексы не поддерживают следующие функции, поддерживаемые в реализации SQL Server XML:
Индексирование узлов со сложными типами XS: типами объединений, типами последовательностей и типами списков.
Индексирование узлов с двоичными типами XS: например, base64Binary и hexBinary.
Указание узлов для индексирования с выражениями XPath, которые содержат подстановочный знак
*в конце. Например,/a/b/c/*,/a//b/*или/a/b/*:c.Индексирование любых осей, отличных от осей дочерних элементов, атрибутов или объектов-потомков. Вариант
//<step>допускается как исключение.Индексирование инструкций и комментариев по обработке XML.
Указание и получение идентификатора для узла с помощью функции id().

Предварительные требования
Следующие предварительные условия должны существовать до создания селективного XML-индекса для XML-столбца в пользовательской таблице.
Кластеризованный индекс должен существовать для первичного ключа таблицы пользователя.
Первичный ключ пользовательской таблицы ограничивается размером в 128 байт при использовании с селективными XML-индексами.
Ключ кластеризации пользовательской таблицы имеет ограничение в 15 столбцов при использовании с селективными XML-индексами.

Ограничения
Общие требования и ограничения
Каждый селективный XML-индекс может быть создан только для одного XML-столбца.
Невозможно создать селективный XML-индекс для столбца, имеющего тип, отличный от XML.
Каждый XML-столбец в таблице может иметь только один селективный XML-индекс.
Каждая таблица может иметь до 249 селективных XML-индексов.
Ограничения для поддерживаемых объектов
Нельзя создавать селективные XML-индексы для следующих объектов:
XML-столбцы в представлении.
Возвращающая табличное значение переменная с XML-столбцами.
Переменные типа XML.
Вычисляемые XML-столбцы.
XML-столбец с глубиной более чем в 128 вложенных узлов.

Ограничения, относящиеся к хранению
Существует ограниченное предельное число узлов из XML-документа, которое можно добавить в индекс. Селективный XML-индекс сопоставляет XML-документы с одной реляционной таблицей. Поэтому он не может содержать более 1024 столбцов со значениями, отличными от NULL, в любой заданной строке таблицы. Кроме того, многие из ограничений для разреженных столбцов также применяются к селективным XML-индексам, поскольку индексы используют разреженные столбцы для хранения.
Максимальное количество столбцов со значениями, отличными от NULL, поддерживаемое в любой заданной строке, зависит от размера данных в столбцах.
В лучшем случае поддерживается 1024 столбца со значениями, отличными от NULL, если все столбцы имеют тип bit.
В худшем случае поддерживается только 236 столбцов со значениями, отличными от NULL, если все столбцы являются большими объектами типа varchar.

Селективные XML-индексы внутренним образом используют от одного до четырех столбцов для каждого индексированного пути узла. Общее количество доступных для индексации узлов лежит в диапазоне от 60 до нескольких сотен узлов и зависит от фактического размера данных в индексированных путях.
В худшем случае, если некоторые или все узлы сопоставлены с помощью
//в определении пути узла, максимальное количество индексируемых узлов равно 60.В лучшем случае, если узлы сопоставлены без использования
//в определении пути узла, максимальное количество индексируемых узлов равно 200.
Селективные XML-индексы перестраиваются при создании и изменении индекса при помощи инструкций CREATE и ALTER.
При создании или изменении селективного XML-индекса при помощи инструкций CREATE и ALTER оно перестраивается в однопотоковом режиме «вне сети». Часто инструкции ALTER отрицательно влияют на производительность запросов к индексированным XML-документам.
Прочие ограничения
Селективные XML-индексы не поддерживаются в указаниях запросов.
Селективные XML-индексы и вторичные селективные XML-индексы не поддерживаются в помощнике по настройке базы данных.
Селективные модуляторы рецепторов андрогена (СМРА)
27 августа 2019 г.
Селективные модуляторы рецепторов андрогена (СМРА) являются новым видом экспериментальных анаболических соединений, использование которых все еще находящится на стадии эксперимента. По сравнению с традиционными анаболическими стероидами они ускоряют рост мышечной массы без возможных побочных эффектов. [1] СМРА связываются с рецепторами андрогена в тканях, изменяя их состав и функции. Существует ряд различных соединений СМРА, которые отличаются друг от друга своим химическим составом. Аналогичным образом механизмы связывания с андрогенными рецепторами зависят от самого соединения. [1] Эффекты СМРА аналогичны таковым у тестостерона и анаболических стероидов. По этой причине они используются в медицине для лечения некоторых заболеваний [2]. В сфере состязательных видов спорта соединения СМРА запрещены с 2008 г. [5].
По этой причине они используются в медицине для лечения некоторых заболеваний [2]. В сфере состязательных видов спорта соединения СМРА запрещены с 2008 г. [5].
СМРА делятся на стероидные и нестероидные соединения. Стероидные СМРА были разработаны на основе химической структуры молекулы тестостерона. [3] Однако большинство разработанных на данный момент СМРА являются нестероидными соединениями [4]. Нестероидные СМРА были изначально обнаружены в результате исследований в университете Теннесси, после чего из этих СМРА были разработаны несколько разнообразных структурных классов [3]. К известным нестероидным СМРА относятся следующие препараты: Энобосарм (остарин), Лигандрол (LGD-4033), Тестолон (RAD-140) и Андарин (GTX-007 или S-4).
Механизмы действия СМРА до сих пор в значительной степени неизвестны, так как их исследования начались лишь недавно. Некоторые механизмы, подтвержденные для активности тканей СМРЭ, также действуют и в отношении СМРА. Они способны активировать рецепторы андрогенов в избранных тканях. Иными словами, они действуют в качестве селективных агонистов для тканей. На практике они применяются для получения эффектов андрогенов с уменьшением их побочных эффектов. Традиционное лечение с применением андрогенов несет в себе риск, например, для сердца, печени и предстательной железы. Предполагается, что СМРА способны активировать рецепторы андрогенов в мышцах и костях при минимальных побочных эффектах. Однако исследования по безопасности и фактической эффективности СМРА пока недостаточны. [4]
Некоторые механизмы, подтвержденные для активности тканей СМРЭ, также действуют и в отношении СМРА. Они способны активировать рецепторы андрогенов в избранных тканях. Иными словами, они действуют в качестве селективных агонистов для тканей. На практике они применяются для получения эффектов андрогенов с уменьшением их побочных эффектов. Традиционное лечение с применением андрогенов несет в себе риск, например, для сердца, печени и предстательной железы. Предполагается, что СМРА способны активировать рецепторы андрогенов в мышцах и костях при минимальных побочных эффектах. Однако исследования по безопасности и фактической эффективности СМРА пока недостаточны. [4]
Применение СМРА и изучение их эффективности происходит в отношении лечения остеопороза, рака предстательной железы, рака груди, гипогонадизма, кахексии, синдрома Альцгеймера, гиперплазии предстательной железы и половой дисфункции. [1, 4] Разработка СМРА также нацелена на устранение вирилизирующих эффектов андрогенов, в связи с чем вышеуказанные заболевания можно лечить, в частности, у женщин без проявления вирилизирующих эффектов, связанных с андрогенами. Тем не менее основным способом лечения в настоящее время является применение тестостерона. [4]
Тем не менее основным способом лечения в настоящее время является применение тестостерона. [4]
Применение в качестве допинга
Анаболические эффекты СМРА и потенциально меньший риск побочных эффектов делают их привлекательными для приверженцев здорового образа жизни и спортсменов, которым необходимо нарастить мышечную массу или силу. В зарубежных интернет-магазинах продаются разнообразные СМРА и пищевые добавки с содержанием СМРА или химически близких к ним веществ. [1] Оценка безопасности СМРА еще только начинается, однако фармацевтические препараты, поставляемые из-за границы, могут содержать примеси, что повышает риски при их применении.
Побочные эффекты недостаточно известны, так как на данный момент было проведено лишь несколько исследований побочных эффектов СМРА в отношении людей. Например, СМРЭ изучаются уже в течение десятилетий, поэтому их побочные эффекты известны лучше. [4]
[4]
Лигандрол (LGD-4033) и Энобосарм в терапевтических дозах прошли успешные испытания на людях. Был обнаружен рост мышечной массы без побочных эффектов, которые характерны для андрогенов. [6, 7] В виде побочных эффектов проявились: экзема, увеличение креатинфосфокиназы после нагрузок, головная боль и ксеростомия. Кроме того, тестовая группа, принимавшая Лигандрол, показала большее число инфекций верхних дыхательных путей, но это обстоятельство не учитывалось как связанное с лекарственным средством. [7]
Так как на данный момент проведено лишь несколько исследований на людях, невозможно гарантировать безопасность и эффективность СМРА. Недавно Европейское агентство лекарственных средств (EMA) отказало в регистрации анаморелина из-за его незначительного эффекта в отношении увеличения мышечной массы. В это же время Управление по санитарному надзору за качеством пищевых продуктов и медикаментов (FDA) США выпустило предостережение относительно рисков, связанных с СМРА, к которым отнесли гепатоксичность и различные проблемы с сердцем. Длительное воздействие данных веществ на людей пока не известно. [8] Тем не менее СМРА выглядит перспективным средством лечения многих заболеваний. В этой связи их необходимо исследовать далее.
Длительное воздействие данных веществ на людей пока не известно. [8] Тем не менее СМРА выглядит перспективным средством лечения многих заболеваний. В этой связи их необходимо исследовать далее.
Вилле Лескинен (Ville Leskinen)
Магистр медицинских наук (MHSc), Dopinglinkki
Saarland University Medical Center — Селективная внутренняя радиотерапия (SIRT)
Selektive interne Radiotherapie (SIRT)
Селективная внутренняя радиотерапия (SIRT)
Метод внутренней селективной терапии SIRT- современный и эффективный вид терапии для лечения опухоли печени, либо метастазов в печени, которые больше не подлежат оперативному методу лечения . SIRT проводится совместно с врачом ядерной медицины и врачом онкологом.
Как проводится SIRT?
Под рентгеновским контролем, через паховую область вводится катетер в печёночную артерию, расположенную вблизи с опухолью. Одновременно через который вводится радиоактивная субстанция, прикреплённая на хорошо переносимыми тканью полимерными шариками (SIR-Spheres®) либо стеклянными шариками (TheraSpheres®), которые вводятся непосредственно в снабжающие опухолью сосуды. Шарики имеют очень маленький диаметр примерно 20-40 мкм это способствует оптимальному заполнению мелких сосудов, снабжающих опухоль кровью. Помеченная на шариках радиоактивная субстанция называется Yttrium (Y-90) -Иттрий 90, обладающая оптимальными качествами для лечения. Она посылает высокую энергию Бета -излучение и тем самым достигает в опухоли высокую лучевую дозу. Так как это излучение обладает незначительной дальностью действия <1 см в ткани, расположенная вокруг опухоли здоровая ткань печени, а также другие близ лежащие органы при проведении SIRT остаются без повреждений. После проведения лечения необходимо в течении 48 часов остаться под стационарным наблюдением, в специализированном для лучевой терапии центре, Клинике лучевой терапии RN- 01.
Одновременно через который вводится радиоактивная субстанция, прикреплённая на хорошо переносимыми тканью полимерными шариками (SIR-Spheres®) либо стеклянными шариками (TheraSpheres®), которые вводятся непосредственно в снабжающие опухолью сосуды. Шарики имеют очень маленький диаметр примерно 20-40 мкм это способствует оптимальному заполнению мелких сосудов, снабжающих опухоль кровью. Помеченная на шариках радиоактивная субстанция называется Yttrium (Y-90) -Иттрий 90, обладающая оптимальными качествами для лечения. Она посылает высокую энергию Бета -излучение и тем самым достигает в опухоли высокую лучевую дозу. Так как это излучение обладает незначительной дальностью действия <1 см в ткани, расположенная вокруг опухоли здоровая ткань печени, а также другие близ лежащие органы при проведении SIRT остаются без повреждений. После проведения лечения необходимо в течении 48 часов остаться под стационарным наблюдением, в специализированном для лучевой терапии центре, Клинике лучевой терапии RN- 01.
Кто назначает SIRT?
Назначение для SIRT проводится совместно с онкологом, медиком нуклеарной медицины и врачом радиологом.
Какая подготовка необходима для проведения SIRT ?
Необходимость проведения SIRT-Терапии решается на врачебном уровне. К тому же, с пациентом проводится подробная беседа о проведении терапии и обязательном предварительном обследовании в отделении.
Подготовка к SIRT-Терапии содержит следующие пункты:
- Подробная история болезни и проводимые до этого момента методы лечения
- Актуальное исследование крови (параметры функции печени, общий анализ крови, опухолевый маркер)
- Проведение магнитно-резонансной томографии /компьютерной томографии печени
- Проведение позитронно-эмиссионной томографии (ПЭТ), для исключения других метастазов вне печени
- Проведение диагностической ангиографии (проверяется сосудистая проходимость артерий печени) с вероятностью окклюзией отходящих сосудов
- Применение инъекции слабого радиоактивного пробного вещества (99mTc-МАА) с последующей планарной сцинтиграфией для проверки окклюзии сосудов, выполненной с помощью ангиографии.

Какие существуют противопоказания для SIRT-Терапии?
Существуют относительные и абсолютные противопоказания, которые различаются в зависимости от выбора радиоактивного препарата для лечения.
Относительные:
- Наличие небольших метастазов вне печени
- Объем коллотерального кровообращения между печенью и легкими между 10 и 20%
- Частичное закрытие воротной вены (vena Portae)
Абсолютные:
- Большие опухолевые поражения вне печени
- Явно нарушенная функция печени с наличием асцита
- Объем кровообращения между печенью и легкими>20%
- Полное закрытие воротной вены (vena Portae)
Какие осложнения или побочные эффекты SIRT известны ?
Осложнения после и во время терапии SIRT очень редки. Как одно из возможных осложнений возможна вероятность имплантации радиоактивных шариков вне печении, к примеру в желудке, или в стенке кишечника. Это может привести к воспалению слизистой оболочке желудка , либо даже к язве желудка. В случае попадания шарика в коллатеральное кровообращение между печенью и легкими может возникнуть радиационная пневмония. Этот вид кровообращения проверяется с помощью пробной субстанции до проведения SIRT. Аллергическая реакция на шарики до сегодняшнего дня не была описана.
Как одно из возможных осложнений возможна вероятность имплантации радиоактивных шариков вне печении, к примеру в желудке, или в стенке кишечника. Это может привести к воспалению слизистой оболочке желудка , либо даже к язве желудка. В случае попадания шарика в коллатеральное кровообращение между печенью и легкими может возникнуть радиационная пневмония. Этот вид кровообращения проверяется с помощью пробной субстанции до проведения SIRT. Аллергическая реакция на шарики до сегодняшнего дня не была описана.
Как проводится SIRT?
SIRT включает в себя следующие этапы :
- Проверка показаний лечащим врачём
- Подробная беседа с пациентом
- Проведение подготовки к обследованию и при необходимости проведение радиологического исследования печени а также сосудов опухоли печени
- Оформление на поступление на стационарный режим, проведение сосудистого статуса печёночных артерий (ангиографии). Для этого применяются заряженные шарики- Иттрий (Yttrium)
 Для этого применяются заряженные шарики- Иттрий (Yttrium)
Для этого применяются заряженные шарики- Иттрий (Yttrium)
- Контролируется распределение шариков с помощью сцинтиграфии
- 48 часовая госпитализация в специализированном для этого отделении RN-01
Требуется ли последующие наблюдение после SIRT?
Последующие наблюдение после SIRT имеет большое значение. Оно проводится совместно с доктором, который направляет и доктором гастроэнтерологом. Оно включает в себя регулярные проверки лабораторных данных функции печени, а также КТ /МРТ, для контроля оценки успеха терапии.
Как часто проводится SIRT?
Как правило один раз, в один день. В единичных случаях при новых изменениях в печени возможно повторное проведение SIRT.
Retromer выполняет селективную функцию сортировки грузов с помощью транспортных носителей для эндосом
Рисунок 1.
Ультраструктурное изменение лизосомальных структур…
Рисунок 1.
Ультраструктурные изменения лизосомных структур и повышенная аутофагия в клетках Vps35 KO.(А)…
Рисунок 1. Ультраструктурные изменения лизосомных структур и повышенная аутофагия в клетках Vps35 KO. (A) Создание CRISPR / Cas9-опосредованных клеток HeLa Vps35 KO и спасательных клеток Vps35-GFP. Равные количества клеточных лизатов из спасательных клеток HeLa, Vps35 KO и Vps35-GFP подвергали SDS-PAGE и иммуноблотировали с антителами против Vps35, Vps26A, Vps29 и тубулина. (B) Электронные микрофотографии спасательных клеток HeLa, Vps35 KO и Vps35-GFP.Увеличенные круглые структуры обозначаются как поздние эндосомные / лизосомные структуры. Масштабные линейки, 2000 нм; на увеличенных изображениях — 500 нм. График представляет процентную объемную плотность лизосомных компартментов относительно цитоплазмы в спасательных клетках HeLa, Vps35 KO и Vps35-GFP (среднее значение ± SEM). Двусторонний критерий Стьюдента t использовали для определения статистической значимости. **, P <0,01; ***, P <0,001. n = два независимых эксперимента по 10 изображений в каждом. (C) Проточный цитометрический анализ подкисления клеток на основе флуоресценции LysoTracker в клетках HeLa и Vps35 KO.График представляет собой среднюю интенсивность флуоресценции внутри клеток HeLa и Vps35 KO (среднее значение ± стандартная ошибка среднего). Двусторонний критерий Стьюдента t был использован для определения статистической значимости ( n = 3). (D) Спасательные клетки HeLa, Vps35 KO и Vps35-GFP фиксировали и коиммуно-метили антителами против LC3-II и LAMP1, а затем флуоресцентными вторичными антителами, конъюгированными с Alexa Fluor.
Масштабные линейки, 2000 нм; на увеличенных изображениях — 500 нм. График представляет процентную объемную плотность лизосомных компартментов относительно цитоплазмы в спасательных клетках HeLa, Vps35 KO и Vps35-GFP (среднее значение ± SEM). Двусторонний критерий Стьюдента t использовали для определения статистической значимости. **, P <0,01; ***, P <0,001. n = два независимых эксперимента по 10 изображений в каждом. (C) Проточный цитометрический анализ подкисления клеток на основе флуоресценции LysoTracker в клетках HeLa и Vps35 KO.График представляет собой среднюю интенсивность флуоресценции внутри клеток HeLa и Vps35 KO (среднее значение ± стандартная ошибка среднего). Двусторонний критерий Стьюдента t был использован для определения статистической значимости ( n = 3). (D) Спасательные клетки HeLa, Vps35 KO и Vps35-GFP фиксировали и коиммуно-метили антителами против LC3-II и LAMP1, а затем флуоресцентными вторичными антителами, конъюгированными с Alexa Fluor. Масштабные линейки 5 мкм. Совместную локализацию между LC3-II и LAMP1 количественно оценивали с помощью коэффициента корреляции Пирсона (среднее значение ± SEM).Двусторонний тест Стьюдента t использовали для определения статистической значимости среди спасательных клеток HeLa, Vps35 KO и Vps35-GFP после аминокислотной стимуляции ( n = 3). ***, P <0,001; ****, P <0,0001. (E) Клетки HeLa и Vps35 KO обрабатывали хлорохином (CQ, 50 мкМ) в течение 6 часов. Клетки собирали, и равные количества образцов белка использовали для SDS-PAGE и иммуноблоттинга с антителами против LC3-II, Vps35 и GAPDH. График представляет уровень LC3-II, нормализованный к GAPDH (среднее значение ± стандартная ошибка среднего).Двусторонний критерий Стьюдента t был использован для определения статистической значимости ( n = 3). *, P <0,05; **, P <0,01. (F) голодные по аминокислотам спасательные клетки HeLa, Vps35 KO и Vps35-GFP обрабатывали 2-кратным раствором незаменимых аминокислот в течение 30 мин, фиксировали ледяным метанолом и коиммуномечены антителами против mTORC1 и LAMP1, после чего следовали от Alexa Fluor — конъюгированные флуоресцентные вторичные антитела (среднее значение ± стандартная ошибка среднего).
Масштабные линейки 5 мкм. Совместную локализацию между LC3-II и LAMP1 количественно оценивали с помощью коэффициента корреляции Пирсона (среднее значение ± SEM).Двусторонний тест Стьюдента t использовали для определения статистической значимости среди спасательных клеток HeLa, Vps35 KO и Vps35-GFP после аминокислотной стимуляции ( n = 3). ***, P <0,001; ****, P <0,0001. (E) Клетки HeLa и Vps35 KO обрабатывали хлорохином (CQ, 50 мкМ) в течение 6 часов. Клетки собирали, и равные количества образцов белка использовали для SDS-PAGE и иммуноблоттинга с антителами против LC3-II, Vps35 и GAPDH. График представляет уровень LC3-II, нормализованный к GAPDH (среднее значение ± стандартная ошибка среднего).Двусторонний критерий Стьюдента t был использован для определения статистической значимости ( n = 3). *, P <0,05; **, P <0,01. (F) голодные по аминокислотам спасательные клетки HeLa, Vps35 KO и Vps35-GFP обрабатывали 2-кратным раствором незаменимых аминокислот в течение 30 мин, фиксировали ледяным метанолом и коиммуномечены антителами против mTORC1 и LAMP1, после чего следовали от Alexa Fluor — конъюгированные флуоресцентные вторичные антитела (среднее значение ± стандартная ошибка среднего). Масштабные линейки 5 мкм. Совместную локализацию mTORC1 с LAMP1 количественно оценивали с помощью коэффициента корреляции Пирсона.Двусторонний тест Стьюдента t показывает разницу между клетками HeLa и Vps35 KO при аминокислотной стимуляции ( n = 3). ***, P <0,001; ****, P <0,0001. (G) Клетки HeLa и Vps35 KO обрабатывали AZD8055 (1 мкМ) в течение 25 часов перед тем, как подвергать SDS-PAGE и иммуноблоттингу с антителами против LC3-II, Vps35 и GAPDH. График представляет уровень экспрессии LC3-II, нормализованный к GAPDH (среднее значение ± стандартная ошибка среднего). Двусторонний критерий Стьюдента t был использован для определения статистической значимости ( n = 3).*, P <0,05.
Масштабные линейки 5 мкм. Совместную локализацию mTORC1 с LAMP1 количественно оценивали с помощью коэффициента корреляции Пирсона.Двусторонний тест Стьюдента t показывает разницу между клетками HeLa и Vps35 KO при аминокислотной стимуляции ( n = 3). ***, P <0,001; ****, P <0,0001. (G) Клетки HeLa и Vps35 KO обрабатывали AZD8055 (1 мкМ) в течение 25 часов перед тем, как подвергать SDS-PAGE и иммуноблоттингу с антителами против LC3-II, Vps35 и GAPDH. График представляет уровень экспрессии LC3-II, нормализованный к GAPDH (среднее значение ± стандартная ошибка среднего). Двусторонний критерий Стьюдента t был использован для определения статистической значимости ( n = 3).*, P <0,05.
Избирательная роль комплекса транслин / trax РНКаза в синаптической пластичности гиппокампа | Молекулярный мозг
Баффингтон С.А., Хуанг В., Коста-Маттиоли М. Трансляционный контроль синаптической пластичности и когнитивной дисфункции. Annu Rev Neurosci. 2014. 37 (1): 17–38.
Annu Rev Neurosci. 2014. 37 (1): 17–38.
CAS PubMed PubMed Central Google ученый
Мартин К.С., Барад М., Кандел ER. Локальный синтез белка и его роль в синапс-специфической пластичности.Curr Opin Neurobiol. 2000. 10 (5): 587–92.
CAS PubMed Google ученый
Саттон М.А., Шуман Э.М. Синтез дендритных белков, синаптическая пластичность и память. Клетка. 2006. 127 (1): 49–58.
CAS PubMed Google ученый
Освальд Стюард Дж. Д., Шеннон Ф. Локальный синтез белка в синапсах. В: Пикель М.С., редактор. Синапс: структура и функции.Лондон: Academic Press; 2014. с. 173–4.
Google ученый
Хафнер А.С., Донлин-Асп П.Г., Лейтч Б., Херцог Э., Шуман Э.М. Локальный синтез белка является повсеместным свойством пре- и постсинаптических компартментов нейронов. Наука. 2019; 364: 6441.
Наука. 2019; 364: 6441.
Google ученый
Фернандес-Моя С.М., Бауэр К.Е., Киблер М.А. Встречайте игроков: локальный перевод в синапсе.Front Mol Neurosci. 2014; 7: 84.
PubMed PubMed Central Google ученый
Thelen MP, Kye MJ. Роль РНК-связывающих белков для локальной трансляции мРНК: последствия при неврологических расстройствах. Front Mol Biosci. 2020; 6: 161.
PubMed PubMed Central Google ученый
Ule J, Darnell RB. РНК-связывающие белки и регуляция синаптической пластичности нейронов.Curr Opin Neurobiol. 2006. 16 (1): 102–10.
CAS PubMed Google ученый
Claußen M, Koch R, Jin Z-Y, Suter B. Функциональная характеристика дрозофилы Translin и Trax. Генетика. 2006. 174 (3): 1337–47.
PubMed PubMed Central Google ученый

Fu X, Shah A, Baraban JM. Быстрое обращение трансляционного молчания: новая роль путей деградации микроРНК в пластичности нейронов.Neurobiol Learn Mem. 2016; 133: 225–32.
CAS PubMed PubMed Central Google ученый
Джендлинг А. Биологическая роль транслина и транслин-ассоциированного фактора-X: метаболизм РНК выходит на первый план. Биохим Дж. 2010; 429 (2): 225–34.
CAS PubMed Google ученый
Li Z, Wu Y, Baraban JM. Комплекс связывания РНК Translin / Trax: ключи к работе в нервной системе.Biochimica et Biophysica Acta (BBA). 2008. 1779 (8): 479–85.
CAS Google ученый
Asada K, Canestrari E, Fu X, Li Z, Makowski E, Wu YC, et al. Спасение дефектов дайсера путем ингибирования нуклеазы, препятствующей образованию кубиков. Cell Rep.2014; 9 (4): 1471–81.
CAS PubMed PubMed Central Google ученый

Барабан Дж. М., Шах А., Фу Х. Множественные пути опосредуют деградацию микроРНК: основное внимание уделяется комплексу РНКазы Translin / Trax.Adv Pharmacol. 2018; 82: 1–20.
CAS PubMed Google ученый
Park AJ, Havekes R, Fu X, Hansen R, Tudor JC, Peixoto L, et al. Обучение побуждает комплекс транслин / trax РНКаз экспрессировать рецепторы активина для постоянной памяти. Элиф. 2017; 6: e27876.
Google ученый
Коста-Маттиоли М., Соссин В.С., Кланн Э., Соненберг Н. Трансляционный контроль длительной синаптической пластичности и памяти.Нейрон. 2009. 61 (1): 10–26.
CAS PubMed PubMed Central Google ученый
Dell’Acqua ML, Smith KE, Gorski JA, Horne EA, Gibson ES, Gomez LL. Регуляция нейрональной передачи сигналов PKA посредством динамики нацеливания AKAP. Eur J Cell Biol. 2006. 85 (7): 627–33.
2006. 85 (7): 627–33.
PubMed Google ученый
Эрнандес П.Дж., Абель Т. Роль синтеза белка в консолидации памяти: прогресс на фоне десятилетий споров.Neurobiol Learn Mem. 2008. 89 (3): 293–311.
CAS PubMed Google ученый
Park AJ, Havekes R, Choi JH, Luczak V, Nie T, Huang T. и др. Пресинаптическая роль PKA в синаптическом тегировании и памяти. Neurobiol Learn Mem. 2014; 114: 101–12.
CAS PubMed PubMed Central Google ученый
Сандерсон Дж. Л., Делл’Аква М.Л. Сигнальные комплексы AKAP в регуляции возбуждающей синаптической пластичности.Невролог. 2011. 17 (3): 321–36.
CAS PubMed PubMed Central Google ученый
Chennathukuzhi V, Stein JM, Abel T., Donlon S, Yang S, Miller JP, et al. Мыши, дефицитные по РНК-связывающему белку яичка и головного мозга, демонстрируют координированную потерю TRAX, снижение фертильности, измененную экспрессию генов в головном мозге и изменения в поведении. Mol Cell Biol. 2003. 23 (18): 6419–34.
Mol Cell Biol. 2003. 23 (18): 6419–34.
CAS PubMed PubMed Central Google ученый
Stein JM, Bergman W, Fang Y, Davison L, Brensinger C, Robinson MB, et al. Поведенческие и нейрохимические изменения у мышей, лишенных РНК-связывающего белка транслина. J Neurosci. 2006. 26 (8): 2184–96.
CAS PubMed PubMed Central Google ученый
Citri A, Soler-Llavina G, Bhattacharyya S, Malenka RC. Долговременная депрессия, зависимая от рецептора N-метил-d-аспартата и метаботропного рецептора глутамата, по-разному регулируется системой убиквитин-протеасома.Eur J Neurosci. 2009. 30 (8): 1443–50.
PubMed PubMed Central Google ученый
Дудек С.М., Медведь М.Ф. Гомосинаптическая долговременная депрессия в области CA1 гиппокампа и эффекты блокады рецептора N-метил-d-аспартата. Proc Natl Acad Sci USA. 1992. 89 (10): 4363–7.
1992. 89 (10): 4363–7.
CAS PubMed PubMed Central Google ученый
Mulkey RM, Malenka RC.Механизмы, лежащие в основе индукции гомосинаптической долговременной депрессии в области CA1 гиппокампа. Нейрон. 1992; 9 (5): 967–75.
CAS PubMed Google ученый
Vecsey CG, Peixoto L, Choi JH, Wimmer M, Jaganath D, Hernandez PJ, et al. Геномный анализ депривации сна выявляет регуляцию трансляции в гиппокампе. Physiol Genomics. 2012; 44 (20): 981–91.
CAS PubMed PubMed Central Google ученый
Финкенштадт PM, Кан WS, Jeon M, Taira E, Tang W., Baraban JM. Соматодендритная локализация Translin, компонента комплекса связывания Translin / Trax РНК. J Neurochem. 2000. 75 (4): 1754–62.
CAS PubMed Google ученый

Даффи С.Н., Нгуен П.В. Постсинаптическое применение пептидного ингибитора цАМФ-зависимой протеинкиназы блокирует экспрессию длительного синаптического потенцирования в нейронах гиппокампа. J Neurosci.2003. 23 (4): 1142–50.
CAS PubMed PubMed Central Google ученый
Ким М., Хуанг Т., Абель Т., Блэквелл К.Т. Временная чувствительность активации протеинкиназы А в поздней фазе долгосрочной потенциации. PLoS Comput Biol. 2010; 6 (2): e1000691.
PubMed PubMed Central Google ученый
Ни Т., Макдоноу, CB, Хуанг Т., Нгуен П.В., Абель Т.Генетическое нарушение закрепления протеинкиназы A показывает роль компартментализованной передачи сигналов киназы в долговременной потенциации тета-всплеска и пространственной памяти. J Neurosci. 2007. 27 (38): 10278–88.
CAS PubMed PubMed Central Google ученый

Scharf MT, Woo NH, Lattal KM, Young JZ, Nguyen PV, Abel T. Синтез белка необходим для улучшения долгосрочной потенциации и долговременной памяти с помощью интервальных тренировок.J Neurophysiol. 2002. 87 (6): 2770–7.
CAS PubMed Google ученый
Ву Н.Х., Даффи С.Н., Абель Т., Нгуен П.В. Временной интервал синаптической стимуляции критически модулирует зависимость LTP от циклической AMP-зависимой протеинкиназы. Гиппокамп. 2003. 13 (2): 293–300.
CAS PubMed Google ученый
Баязитов И.Т., Ричардсон Р.Дж., Фрике Р.Г., Захаренко С.С.Медленные пресинаптические и быстрые постсинаптические компоненты сложной долгосрочной потенциации. J Neurosci. 2007. 27 (43): 11510–21.
CAS PubMed PubMed Central Google ученый
Chavez-Noriega LE, Stevens CF. Повышенное высвобождение медиатора в возбуждающих синапсах, вызванное прямой активацией аденилатциклазы в срезах гиппокампа крысы.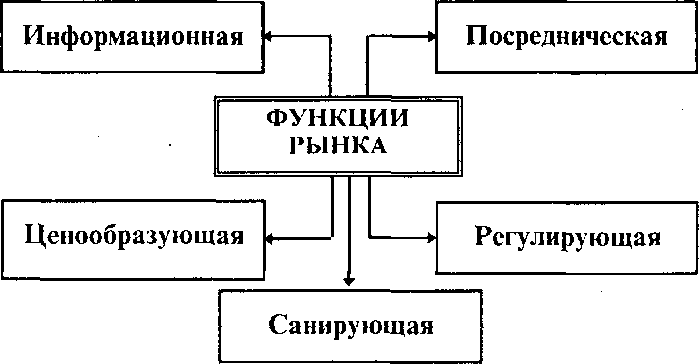 J Neurosci. 1994. 14 (1): 310–7.
J Neurosci. 1994. 14 (1): 310–7.
CAS PubMed PubMed Central Google ученый
Трюдо Л.Е., Эмери Д.Г., Хейдон П.Г. Прямая модуляция секреторного аппарата лежит в основе PKA-зависимого синаптического облегчения в нейронах гиппокампа. Нейрон. 1996. 17 (4): 789–97.
CAS PubMed Google ученый
Bear MF, Huber KM, Warren ST. Теория mGluR хрупкой умственной отсталости X. Trends Neurosci. 2004. 27 (7): 370–7.
CAS PubMed Google ученый
Хубер К.М., Галлахер С.М., Уоррен С.Т., Медведь М.Ф. Измененная синаптическая пластичность в мышиной модели хрупкой умственной отсталости X. Proc Natl Acad Sci USA. 2002. 99 (11): 7746–50.
CAS PubMed PubMed Central Google ученый
Сидоров М.С., Ауэрбах Б.Д., Медведь М. Ф. Белок умственной отсталости Fragile X и синаптическая пластичность. Мол мозг. 2013; 6:15.
Ф. Белок умственной отсталости Fragile X и синаптическая пластичность. Мол мозг. 2013; 6:15.
CAS PubMed PubMed Central Google ученый
Лу Й, Чжан М., Лим И.А., Холл Д.Д., Аллен М., Медведева Ю. и др. AKAP150-заякоренная активность PKA важна для LTD во время фазы индукции. J Physiol. 2008. 586 (17): 4155–64.
CAS PubMed PubMed Central Google ученый
Сандерсон Дж. Л., Горски Дж. А., Dell’Acqua ML. NMDA рецептор-зависимая LTD требует временного синаптического включения Ca (2) (+) — проницаемых AMPAR, опосредованных AKAP150-заякоренными PKA и кальциневрином.Нейрон. 2016; 89 (5): 1000–15.
CAS PubMed PubMed Central Google ученый
Weng YT, Chien T, Kuan II, Chern Y. Комплекс TRAX, DISC1 и GSK3 при психических расстройствах и терапевтических вмешательствах. J Biomed Sci. 2018; 25 (1): 71.
J Biomed Sci. 2018; 25 (1): 71.
PubMed PubMed Central Google ученый
Фрей У., Моррис РГМ. Синаптическая маркировка и долгосрочное потенцирование.Природа. 1997. 385 (6616): 533–6.
CAS Google ученый
Редондо Р.Л., Моррис Р.Г. Делаем воспоминания долговечными: гипотеза синаптического тегирования и захвата. Nat Rev Neurosci. 2011; 12 (1): 17–30.
CAS PubMed Google ученый
Connor SA, Hoeffer CA, Klann E, Nguyen PV. Белок умственной отсталости Fragile X регулирует гетеросинаптическую пластичность в гиппокампе.Learn Mem. 2011; 18 (4): 207–20.
CAS PubMed PubMed Central Google ученый
Каудерер Б.С., Кандел Э.Р. Захват зависимого от синтеза белка компонента долговременной депрессии. Proc Natl Acad Sci USA. 2000. 97 (24): 13342–7.
2000. 97 (24): 13342–7.
CAS PubMed PubMed Central Google ученый
Манахан-Воган Д., Кулла А., Фрей Дж. Требование трансляции, но не транскрипции для поддержания долговременной депрессии в области CA1 свободно движущихся крыс.J Neurosci. 2000. 20 (22): 8572–6.
CAS PubMed PubMed Central Google ученый
Чан SP, Slack FJ. опосредованное микроРНК молчание внутри Р-телец. RNA Biol. 2006; 3 (3): 97–100.
CAS PubMed Google ученый
Ло Й, На З, Славофф С.А. П-тела: состав, свойства и функции. Биохимия. 2018; 57 (17): 2424–31.
CAS PubMed Google ученый
Райгор Д., Хэнли Дж. Плюсы и минусы миРНК-опосредованного подавления гена во время синаптической пластичности нейронов. Некодирующая РНК. 2016; 2 (1): 1.
2016; 2 (1): 1.
PubMed Central Google ученый
Oh JY, Kwon A, Jo A, Kim H, Goo YS, Lee JA и др. Зависимая от активности синаптическая локализация обрабатывающих тел и их роль в структурной пластичности дендритов. J Cell Sci. 2013; 126 (Pt 9): 2114–23.
CAS PubMed Google ученый
Селективная роль вентромедиального субталамического ядра в ингибирующем контроле
Первоначальная идея о том, что STN является ключевым узлом в мозговой сети ингибирования ответа, возникла в основном из данных нейровизуализации (Aron and Poldrack, 2006; Aron et al., 2007; Ли и др., 2008; Sharp et al., 2010) и исследования поражений (Baunez et al., 1995; Eagle et al., 2008; Obeso et al., 2014; Nishioka et al., 2008). Однако многие аспекты нейрональной передачи сигналов во время подавления действия оставались неясными, несмотря на недавние электрофизиологические записи STN, выполненные у крыс (Schmidt et al. , 2013), обезьян (Isoda and Hikosaka, 2008) и людей (Bastin et al., 2014; Benis et al. ., 2014; Ray et al., 2012; Benis et al., 2016). Здесь мы обнаружили, что нейроны, расположенные в наиболее передней и вентромедиальной части STN, демонстрируют фазовое изменение активности, подходящее для активной роли в остановке.Наша новая парадигма позволила нам подтвердить, что некоторая активность STN является стоп-селективной, а не отражает множество потенциально смешанных факторов. Насколько нам известно, эти результаты являются первым доказательством избирательного кодирования остановки на клеточном уровне в STN здоровых приматов.
, 2013), обезьян (Isoda and Hikosaka, 2008) и людей (Bastin et al., 2014; Benis et al. ., 2014; Ray et al., 2012; Benis et al., 2016). Здесь мы обнаружили, что нейроны, расположенные в наиболее передней и вентромедиальной части STN, демонстрируют фазовое изменение активности, подходящее для активной роли в остановке.Наша новая парадигма позволила нам подтвердить, что некоторая активность STN является стоп-селективной, а не отражает множество потенциально смешанных факторов. Насколько нам известно, эти результаты являются первым доказательством избирательного кодирования остановки на клеточном уровне в STN здоровых приматов.
Задача стоп-сигнала широко использовалась для исследования нервного субстрата подавления ответа глазодвигательного аппарата (Hanes et al., 1998; Paré and Hanes, 2003; Ito et al., 2003; Stuphorn et al., 2000) и скелетомоторной (Bastin et al., 2014; Scangos, Stuphorn, 2010; Mirabella et al., 2011; Benis et al., 2016; Chen et al., 2010) системы. Эта парадигма накладывает жесткие временные ограничения в том смысле, что нейронный отклик должен произойти в рассчитанном SSRT, чтобы он способствовал остановке. Более одной трети наших нейронов STN соответствовали этому временному критерию (рис. 3–4). Среди этих клеток наиболее распространенным паттерном реакции была активация, связанная с движением, которая быстро затухала в ответ на стоп-сигнал, аналогично активности, наблюдаемой в мышцах-агонистах (рис. 5А).Примечательно, что второй паттерн реакции состоял из фазового увеличения разряда в ответ на стоп-сигнал. Сосуществование этих двух паттернов отклика согласуется с ролями STN как в выполнении движения, так и в остановке движения (более подробно обсуждается ниже). Модель гонки в своей простейшей форме предсказывает, что активность, связанная с остановкой, должна предшествовать или, по крайней мере, совпадать с усеченным наращиванием активности, связанной с движением. Однако мы обнаружили, что среднее время нейтрализации для клеток движения было немного раньше, чем для стоп-клеток.Более того, связанная с остановкой активность предшествовала самому раннему вызванному остановкой усечению ЭМГ-активности всего на 10 мс.
Более одной трети наших нейронов STN соответствовали этому временному критерию (рис. 3–4). Среди этих клеток наиболее распространенным паттерном реакции была активация, связанная с движением, которая быстро затухала в ответ на стоп-сигнал, аналогично активности, наблюдаемой в мышцах-агонистах (рис. 5А).Примечательно, что второй паттерн реакции состоял из фазового увеличения разряда в ответ на стоп-сигнал. Сосуществование этих двух паттернов отклика согласуется с ролями STN как в выполнении движения, так и в остановке движения (более подробно обсуждается ниже). Модель гонки в своей простейшей форме предсказывает, что активность, связанная с остановкой, должна предшествовать или, по крайней мере, совпадать с усеченным наращиванием активности, связанной с движением. Однако мы обнаружили, что среднее время нейтрализации для клеток движения было немного раньше, чем для стоп-клеток.Более того, связанная с остановкой активность предшествовала самому раннему вызванному остановкой усечению ЭМГ-активности всего на 10 мс. Один из возможных выводов состоит в том, что точное положение STN в моделях реактивной остановки может потребовать модификации в зависимости от идентификации областей мозга с более ранними действиями, связанными с остановкой. Альтернативная интерпретация, согласно которой активность, связанная с STN Stop, передается в двигательные центры по быстропроводящему пути, обсуждается ниже.
Один из возможных выводов состоит в том, что точное положение STN в моделях реактивной остановки может потребовать модификации в зависимости от идентификации областей мозга с более ранними действиями, связанными с остановкой. Альтернативная интерпретация, согласно которой активность, связанная с STN Stop, передается в двигательные центры по быстропроводящему пути, обсуждается ниже.
Классическая задача стоп-сигнала, когда использовалась сама по себе, как и в предыдущих нейрофизиологических исследованиях (Aron and Poldrack, 2006; Schmidt et al., 2013; Scangos and Stuphorn, 2010), может объединить несколько различных когнитивных процессов (Duann et al., 2009; Erika-Florence et al., 2014; Hampshire et al., 2010). Во-первых, поскольку стоп-сигнал является нечастым (15–30% испытаний) и поведенчески значимым, некоторые нейронные реакции на стоп-сигнал могут отражать обработку вниманием неожиданного визуального сигнала. Действительно, STN является частью сети, в которой реализованы визуальное внимание и значимость событий (Baunez and Robbins, 1997; Baunez et al. , 2007; Бочкова и др., 2011; Schmalbach et al., 2014). Во-вторых, выполнение задачи с сигналом остановки требует нескольких аспектов исполнительного контроля, включая изменения когнитивного набора, усиление сенсорной реакции, содействие желаемой реакции и мониторинг производительности. Таким образом, некоторые фазовые ответы на стоп-сигнал могут отражать подпроцессы, связанные с переключением.
, 2007; Бочкова и др., 2011; Schmalbach et al., 2014). Во-вторых, выполнение задачи с сигналом остановки требует нескольких аспектов исполнительного контроля, включая изменения когнитивного набора, усиление сенсорной реакции, содействие желаемой реакции и мониторинг производительности. Таким образом, некоторые фазовые ответы на стоп-сигнал могут отражать подпроцессы, связанные с переключением.
Наша парадигма стоп-сигнала распутала эти факторы, представив идентичный визуальный стимул для подачи неожиданных команд либо для подавления начального действия, либо для инициирования достигаемого движения. Switch-stop и switch-go Типы испытаний были одинаково нечастыми и поведенчески значимыми, и оба требовали поведенческого переключения с привычного на контролируемое реагирование. Сравнение активности STN между двумя типами испытаний переключателей выявило две категории ответов с короткой задержкой (рисунок 6). Хотя стандартные критерии классифицировали бы все как играющие активную роль в остановке, 43% ячеек с короткими откликами на остановку реагировали аналогичным или более сильным образом внезапным инициированием действия в испытаниях switch-go . В полном соответствии с ролью в процессах внимания или переключения, предложенной Исодой и Хикосакой (Isoda and Hikosaka, 2008), эти нейроны STN изменяли свою активность, когда животному приходилось быстро менять поведение от стандартного к контролируемому поведению, требующему большего внимания.
В полном соответствии с ролью в процессах внимания или переключения, предложенной Исодой и Хикосакой (Isoda and Hikosaka, 2008), эти нейроны STN изменяли свою активность, когда животному приходилось быстро менять поведение от стандартного к контролируемому поведению, требующему большего внимания.
Вторая подгруппа нейронов STN активировалась выборочно только тогда, когда животному приходилось отменять текущее действие в испытаниях с переключением-остановкой . Эти клетки не перекрывались ни категориально, ни анатомически с нейронами STN, которые, скорее всего, непосредственно участвовали в активации мышц-антагонистов, а именно те, которые реагировали на проприоцептивную стимуляцию и концентрировались в скелетомоторной территории STN (Nambu et al., 2002; Родитель и Хазрати, 1995; Baron et al., 2002). Вместе эти результаты обеспечивают последовательную поддержку наличия избирательного кодирования реактивного останова в подмножестве нейронов STN после контроля ряда потенциальных затруднений.
В нескольких предыдущих исследованиях изучалась единичная активность STN во время выполнения задачи типа ингибирования ответа. Schmidt et al. (2013) описали популяцию нейронов STN у крысы, которая реагировала с короткой задержкой на сигнал Stop. Примечательно, что, в отличие от настоящего исследования, эти нейроны также примерно с одинаковой задержкой и величиной реагировали на сигнал Go, который был представлен во всех испытаниях (Mallet et al., 2016). Наконец, Schmidt et al. предоставили доказательства того, что стоп-селективная активность проявляется в анатомически ограниченной области ретикулатной черной субстанции (SNr), выходного ядра BG, расположенного ниже STN. Многие факторы различались между исследованием Шмидта и нашим, включая виды, задачи, методы сбора и анализа, любая комбинация которых может объяснить различия между результатами. В недавнем исследовании с участием людей, перенесших операцию DBS по поводу болезни Паркинсона, Benis et al.(2016) описали отдельные субпопуляции нейронов STN, которые избирательно и с короткой задержкой реагировали на сигналы задач Stop и Go , аналогично нашим результатам. Исода и Хикосака (2008) записали данные с STN макак во время задания, которое требовало от животных переключения поведения между знакомыми и неожиданными глазодвигательными реакциями, что в некоторой степени похоже на наши испытания с переключением на режим. Как и мы, они обнаружили отдельную подгруппу нейронов STN, которая избирательно и с короткой задержкой реагировала на неожиданный импульс переключения.Однако ни одно из предыдущих исследований не диссоциировало активности STN, связанных с реактивной остановкой, реактивным переключением и упреждающим (NoGo) контролем ответа, не проверялось на возможное совпадение между активностью, связанной с остановкой, и скелетомоторной реактивностью, или не рассматривалось, сосредоточена ли связанная с остановкой активность в суб- регион СТН.
Исода и Хикосака (2008) записали данные с STN макак во время задания, которое требовало от животных переключения поведения между знакомыми и неожиданными глазодвигательными реакциями, что в некоторой степени похоже на наши испытания с переключением на режим. Как и мы, они обнаружили отдельную подгруппу нейронов STN, которая избирательно и с короткой задержкой реагировала на неожиданный импульс переключения.Однако ни одно из предыдущих исследований не диссоциировало активности STN, связанных с реактивной остановкой, реактивным переключением и упреждающим (NoGo) контролем ответа, не проверялось на возможное совпадение между активностью, связанной с остановкой, и скелетомоторной реактивностью, или не рассматривалось, сосредоточена ли связанная с остановкой активность в суб- регион СТН.
Наконец, аналогичные паттерны реакции, связанной с движением и остановкой, были обнаружены в лобной коре глаза и верхнем холмике макак, выполняющих задачу, запрещающую движение глаз (Schall and Godlove, 2012; Hanes et al. , 1998; Паре и Ханес, 2003). Хотя STN включает глазодвигательную территорию (Matsumura et al., 1992), мы не нашли доказательств того, что указанная здесь активность, связанная с остановкой, действительно отражала движения глаз (Рисунок 6 — рисунок в приложении 1).
, 1998; Паре и Ханес, 2003). Хотя STN включает глазодвигательную территорию (Matsumura et al., 1992), мы не нашли доказательств того, что указанная здесь активность, связанная с остановкой, действительно отражала движения глаз (Рисунок 6 — рисунок в приложении 1).
Использование задачи Go / NoGo (Bari and Robbins, 2013; Aron, 2011; Dalley et al., 2011) также позволило нам изучить участие STN в устойчивом (« проактивном ») поддержании подавляющего движения поведенческого набора между время инструкции NoGo и предъявления триггерного стимула через несколько секунд.Полностью четверть нейронов STN проявляла устойчивую активность преимущественно после сигнала NoGo (рис. 8). Эти клетки не были ограничены определенной территорией STN, что резко контрастировало с анатомически ограниченным расположением стопорных клеток. Распределение ячеек NoGo по STN подразумевает, что многие функциональные подсхемы через STN активируются во время этого поведения. Вероятно, что активность NoGo, как определено здесь, на самом деле отражает множество независимых когнитивных и моторных процессов, анализ которых потребует дальнейшего уточнения поведенческих парадигм. Предыдущие исследования показали, что полосатое тело и «непрямой» путь полосато-паллидного STN участвуют в проактивном контроле (Casey et al., 1997; Kelly et al., 2004; Smittenaar et al., 2013; Vink et al., 2005). Хотя настоящие результаты не рассматривают эту возможность, они утверждают, что проактивное и реактивное подавление движений задействует совершенно разные нейронные механизмы. Мало того, что действия Stop и NoGo были по-разному распределены в пределах STN, но и наличие активности NoGo в отдельном модуле не имело силы для прогнозирования того, будет ли этот модуль также предпочтительно реагировать на сигналы switch-stop или switch-go (Рисунок 8E и Рисунок 8 — дополнение к рисунку 2).
Предыдущие исследования показали, что полосатое тело и «непрямой» путь полосато-паллидного STN участвуют в проактивном контроле (Casey et al., 1997; Kelly et al., 2004; Smittenaar et al., 2013; Vink et al., 2005). Хотя настоящие результаты не рассматривают эту возможность, они утверждают, что проактивное и реактивное подавление движений задействует совершенно разные нейронные механизмы. Мало того, что действия Stop и NoGo были по-разному распределены в пределах STN, но и наличие активности NoGo в отдельном модуле не имело силы для прогнозирования того, будет ли этот модуль также предпочтительно реагировать на сигналы switch-stop или switch-go (Рисунок 8E и Рисунок 8 — дополнение к рисунку 2).
STN получает топографически организованные глутаматергические входы из большинства регионов лобной коры (Nambu et al., 2002; Haynes and Haber, 2013; Monakow et al., 1978). Среди них нижняя лобная кора (IFC; потенциальный гомолог у макак: область 45) была тесно связана с реактивной остановкой на основе поражения (Aron et al. , 2003; Swick et al., 2008) и стимуляции (Verbruggen et al. , 2010; Wessel et al., 2013) исследования. Хотя конкретная топография проекции IFC-to-STN еще не определена, исследования трассировки трактов показывают, что большинство префронтальных областей [i.е., вентромедиальная, орбитофронтальная, передняя поясная извилина и дорсальная префронтальная кора (Haynes and Haber, 2013)] проецируются на вентральную и медиальную части STN, которые охватывают регионы, где располагаются клетки Stop и Switch . Эта часть STN также получает сильные GABAergic входы от рострально-медиального внешнего pallidum (часть ассоциативного контура BG) и вентрального pallidum (лимбического контура BG) (Haynes and Haber, 2013; Shink et al., 1996; Karachi и другие., 2005), и дополнительные глутаматергические входы из парафасцикулярного ядра таламуса, также связанного с лимбической цепью ядра (Sadikot et al., 1992). В соответствии с его афферентной связностью, эта вентромедиальная область содержала несколько клеток с активностью, подобной движению (Figure 4B), делая маловероятным, что процессы Stop и Go сравниваются локально внутри вентромедиального STN.
, 2003; Swick et al., 2008) и стимуляции (Verbruggen et al. , 2010; Wessel et al., 2013) исследования. Хотя конкретная топография проекции IFC-to-STN еще не определена, исследования трассировки трактов показывают, что большинство префронтальных областей [i.е., вентромедиальная, орбитофронтальная, передняя поясная извилина и дорсальная префронтальная кора (Haynes and Haber, 2013)] проецируются на вентральную и медиальную части STN, которые охватывают регионы, где располагаются клетки Stop и Switch . Эта часть STN также получает сильные GABAergic входы от рострально-медиального внешнего pallidum (часть ассоциативного контура BG) и вентрального pallidum (лимбического контура BG) (Haynes and Haber, 2013; Shink et al., 1996; Karachi и другие., 2005), и дополнительные глутаматергические входы из парафасцикулярного ядра таламуса, также связанного с лимбической цепью ядра (Sadikot et al., 1992). В соответствии с его афферентной связностью, эта вентромедиальная область содержала несколько клеток с активностью, подобной движению (Figure 4B), делая маловероятным, что процессы Stop и Go сравниваются локально внутри вентромедиального STN.
Какой эфферентный путь позволяет связанной с остановкой активности в STN подавлять двигательную активность с короткой задержкой? Настоящие результаты не затрагивают этот вопрос напрямую, но они значительно ограничивают жизнеспособные ответы.Общепринятая модель утверждает, что первый эфферентный шаг от STN — это сильно расходящаяся проекция на выходные ядра BG [т.е. внутренний паллидум (GPi) и SNr] (Aron, 2011; Aron and Poldrack, 2006; Schmidt et al. ., 2013). Дивергенция в этом пути считается решающей для трансляции активности, связанной с остановкой, с ассоциативной / лимбической территории STN на ГАМКергические нейроны в скелетомоторной и глазодвигательной территориях GPi и SNr, соответственно. Необходимость дивергенции в какой-то момент цепи подкрепляется доказательствами того, что реактивное останавливающее поведение связано с очевидно глобальным подавлением моторной и даже когнитивной функции (Wessel and Aron, 2017).Однако ограниченные данные подтверждают расхождение в пути STN-GPi / SNr (Hazrati and Parent, 1992), тогда как многочисленные доказательства указывают на то, что вместо этого эта аксональная проекция очень специфична и топографически организована (Shink et al. , 1996; Smith et al. ., 1998; Коломиец и др., 2001; Келли, Стрик, 2004; Миячи и др., 2006). Возможные альтернативные пути включают прямую проекцию STN на тегментальное педункулопонтинное ядро (PPN) (Shink et al., 1996; Hazrati and Parent, 1992; Parent and Smith, 1987; Nakano et al., 1990; Smith et al., 1990), а также дисинаптический путь STN-GPi / SNr-PPN. В этом отношении интересно отметить, что избирательная активация холинергических нейронов вентральной PPN запускает кратковременное и широко распространенное торможение двигательной активности (Takakusaki et al., 1993; Takakusaki et al., 2016; Kohyama et al. , 1998) в дополнение к ряду функций оповещения и модуляции поведения (Mena-Segovia et al., 2004; Gut and Winn, 2016). Дальнейшие исследования необходимы для выяснения степени дивергенции и конкретных участков окончания проекций от вентромедиального STN к PPN и GPi / SNr.И, как упоминалось во введении, вполне вероятно, что реактивная остановка, подобная той, что изучается здесь, зависит от каскада процессов подавления движений и множества мозговых путей (Pouget et al.
, 1996; Smith et al. ., 1998; Коломиец и др., 2001; Келли, Стрик, 2004; Миячи и др., 2006). Возможные альтернативные пути включают прямую проекцию STN на тегментальное педункулопонтинное ядро (PPN) (Shink et al., 1996; Hazrati and Parent, 1992; Parent and Smith, 1987; Nakano et al., 1990; Smith et al., 1990), а также дисинаптический путь STN-GPi / SNr-PPN. В этом отношении интересно отметить, что избирательная активация холинергических нейронов вентральной PPN запускает кратковременное и широко распространенное торможение двигательной активности (Takakusaki et al., 1993; Takakusaki et al., 2016; Kohyama et al. , 1998) в дополнение к ряду функций оповещения и модуляции поведения (Mena-Segovia et al., 2004; Gut and Winn, 2016). Дальнейшие исследования необходимы для выяснения степени дивергенции и конкретных участков окончания проекций от вентромедиального STN к PPN и GPi / SNr.И, как упоминалось во введении, вполне вероятно, что реактивная остановка, подобная той, что изучается здесь, зависит от каскада процессов подавления движений и множества мозговых путей (Pouget et al. , 2017; Schmidt and Berke, 2017).
, 2017; Schmidt and Berke, 2017).
Также стоит обсудить наше частое наблюдение в STN увеличения активности при движении, которое было усечено сразу после сигнала остановки (рис. 4A), активность, названная подобной движению, из-за ее сходства по форме с ЭМГ мышц-агонистов (рис. 5A). ).Осмысление этого типа активности осложняется широко распространенным мнением о том, что STN влияет на центры управления двигателем в первую очередь через прямое возбуждение выходных ГАМКергических нейронов BG (в GPi и SNr) (Nambu et al., 2002; Wichmann et al. др., 1994; Хамада, ДеЛонг, 1992). При передаче через этот путь увеличение перидвижений активности STN должно подавлять движение, а связанное с внезапной остановкой сокращение активности, облегчающее движение — оба эффекта противоположны ролям, предполагаемым для STN в реактивной остановке ( Арон и Полдрак, 2006).Одно из недавно предложенных решений состоит в том, что другой эфферентный путь STN, который ингибирует нейроны, выводящие BG через дисинаптический путь через внешний бледный (GPe), может доминировать над функцией цепи в определенных поведенческих контекстах (Jantz et al. , 2017). Это, однако, не решает основную проблему объяснения одновременного присутствия в STN моделей активности, связанных с Stop и связанных с движением. Ответы на эту загадку могут дать исследования организации проекций из субрегионов STN, активности отдельных подтипов нейронов STN во время выполнения стоп-задания (Sato et al., 2000), или как эта, казалось бы, парадоксальная комбинация активностей STN интерпретируется в нижележащих ядрах.
, 2017). Это, однако, не решает основную проблему объяснения одновременного присутствия в STN моделей активности, связанных с Stop и связанных с движением. Ответы на эту загадку могут дать исследования организации проекций из субрегионов STN, активности отдельных подтипов нейронов STN во время выполнения стоп-задания (Sato et al., 2000), или как эта, казалось бы, парадоксальная комбинация активностей STN интерпретируется в нижележащих ядрах.
Настоящие результаты в целом согласуются с существующими линиями доказательств, которые указывают на участие вентромедиального STN в различных клинических нарушениях ингибиторного контроля. Тематические исследования показали, что DBS в вентральном STN оказывает большее влияние на ингибирующее контролирующее поведение, чем стимуляция в дорсальном STN (Greenhouse et al., 2013; Hershey et al., 2010). Фактически, это анатомическое расположение в настоящее время является целью STN-DBS для лечения тяжелого обсессивно-компульсивного расстройства (Mallet et al. , 2008), что считается нарушением тормозного контроля. И локальные микровливания бикукуллина (антагониста ГАМК) только в ту часть STN, индуцированную стереотипами и гиперактивным поведением (Karachi et al., 2009). Настоящее исследование является первым, показывающим, что единичное кодирование реактивной остановки также ограничивается передним вентромедиальным STN. Важно признать, что тормозящий контроль — это общий термин, охватывающий разрозненный набор поведенческих способностей и подчиненных цепей мозга.Наши результаты подтверждают мнение о том, что вентромедиальный STN является важным узлом в одной из этих цепей мозга, но диапазон ролей, играемых этой цепью в конкретном поведении и поведенческих расстройствах, еще предстоит определить. Интересно отметить, что активность, связанная с переключением быстрого ответа (т.е. Switch клеток), также была ограничена одной и той же анатомической горячей точкой (рис. 7C). Необходимы дальнейшие исследования, чтобы определить, отражают ли описанные здесь действия, связанные с Stop- и Switch , действительно отдельные процессы или, скорее, градуированные смеси сигналов, отражающие различные аспекты реактивного управления.
, 2008), что считается нарушением тормозного контроля. И локальные микровливания бикукуллина (антагониста ГАМК) только в ту часть STN, индуцированную стереотипами и гиперактивным поведением (Karachi et al., 2009). Настоящее исследование является первым, показывающим, что единичное кодирование реактивной остановки также ограничивается передним вентромедиальным STN. Важно признать, что тормозящий контроль — это общий термин, охватывающий разрозненный набор поведенческих способностей и подчиненных цепей мозга.Наши результаты подтверждают мнение о том, что вентромедиальный STN является важным узлом в одной из этих цепей мозга, но диапазон ролей, играемых этой цепью в конкретном поведении и поведенческих расстройствах, еще предстоит определить. Интересно отметить, что активность, связанная с переключением быстрого ответа (т.е. Switch клеток), также была ограничена одной и той же анатомической горячей точкой (рис. 7C). Необходимы дальнейшие исследования, чтобы определить, отражают ли описанные здесь действия, связанные с Stop- и Switch , действительно отдельные процессы или, скорее, градуированные смеси сигналов, отражающие различные аспекты реактивного управления.
Избирательная роль кальциневрина Aα в синаптической депотентации в гиппокампе
Abstract
Фармакологические исследования показали, что для длительной потенциации (LTP), долговременной депрессии (LTD) и депотенции, трех форм синаптической пластичности в гиппокампе, требуется активность фосфатазы кальциневрина. В центральной нервной системе обнаружены по крайней мере две разные изоформы кальциневрина. Чтобы выяснить, требуются ли для всех этих форм синаптической пластичности одни и те же изоформы кальциневрина, мы исследовали LTD, депотенцию и LTP у мышей, лишенных преобладающей изоформы кальциневрина в центральной нервной системе, мышей Aα — / — .Депотенциация была полностью отменена, в то время как ни LTD, ни LTP не пострадали. Эти исследования предоставляют генетические доказательства того, что изоформа кальциневрина Aα важна для обращения LTP в гиппокампе, и указывают на то, что депотенциация и LTD действуют через несколько разные молекулярные механизмы.
Кальциневрин — это серин / треонин-протеиновая фосфатаза, которая широко экспрессируется в центральной нервной системе, включая гиппокамп (1, 2). Из-за своего высокого сродства к комплексу Ca 2+ / кальмодулин, кальциневрин, как полагают, играет критическую роль в синаптической пластичности (3-6).В частности, фармакологические исследования с использованием ингибиторов протеинфосфатазы показали, что как длительная депрессия (LTD), так и депотентация требуют активации протеинфосфатаз (7-9), и вместе с генетическими исследованиями также предположили, что эти протеинфосфатазы противодействуют действию протеина. киназы, необходимые для долговременной потенциации (ДП) (5, 10, 11).
Кальциневрин также может играть непосредственную роль в LTP, поскольку ингибиторы кальциневрина влияют на индукцию и экспрессию LTP (12–14).Поскольку существует по крайней мере две изоформы кальциневрина, эти данные свидетельствуют о том, что разные изоформы могут участвовать в LTD и LTP или что фармакологические ингибиторы вызывают не зависящие от кальциневрина эффекты, которые препятствуют индукции LTD или LTP (15). Чтобы решить эти проблемы, мы исследовали LTD, депотенцию и LTP у мышей, лишенных преобладающей изоформы кальциневрина в центральной нервной системе, Aα. Мы обнаружили, что у мышей с кальциневрином Aα — / — депотенцирование полностью отменялось, в то время как ни LTD, ни LTP не были затронуты.
Чтобы решить эти проблемы, мы исследовали LTD, депотенцию и LTP у мышей, лишенных преобладающей изоформы кальциневрина в центральной нервной системе, Aα. Мы обнаружили, что у мышей с кальциневрином Aα — / — депотенцирование полностью отменялось, в то время как ни LTD, ни LTP не были затронуты.
МАТЕРИАЛЫ И МЕТОДЫ
Иммуногистохимия.
Для определения локализации изоформ кальциневрина A в центральной нервной системе использовали кроличьи поликлональные антипептидные антисыворотки к изоформе кальциневрина Aα (пептид LSNSSNIQ; R2929) и к изоформе кальциневрина Aβ (LMTEGEDEFDG; R2948) для окрашивания парафиновых срезов тканей мозга. от пяти мышей дикого типа и пяти мутантных мышей. Чтобы идентифицировать возможные дополнительные тонкие аномалии у мутантов, кроличьи антитела к глиальному фибриллярному кислому белку (предоставлены L.F. Eng, Стэнфордский университет), синаптофизин (Zymed) и синапсин 1 (Biogenesis, Sandown, NH) также использовались для иммуногистохимических исследований. Парафиновые срезы толщиной шесть микрометров окрашивали с использованием наборов для окрашивания иммунопероксидазой (Vector Laboratories), как описано (30). Вкратце, после депарафинизации и гидратации предметные стекла промывали PBS (pH 7,3) и последовательно инкубировали в 10% нормальной козьей сыворотке в PBS, первичных антителах (разведенных 1: 100–1: 200 в PBS), 0,03% H 2 O 2 в PBS, биотинилированном антикроличьем Ig, и авидин-биотиновый комплекс пероксидазы хрена.Срезы промывали PBS между каждым этапом инкубации. Продукт иммунопероксидазной реакции визуализировали с помощью 3-амино-9-этилкарбазола (Aldrich) и фиксировали в формол-ацетате. Срезы контрастировали гематоксилином.
Парафиновые срезы толщиной шесть микрометров окрашивали с использованием наборов для окрашивания иммунопероксидазой (Vector Laboratories), как описано (30). Вкратце, после депарафинизации и гидратации предметные стекла промывали PBS (pH 7,3) и последовательно инкубировали в 10% нормальной козьей сыворотке в PBS, первичных антителах (разведенных 1: 100–1: 200 в PBS), 0,03% H 2 O 2 в PBS, биотинилированном антикроличьем Ig, и авидин-биотиновый комплекс пероксидазы хрена.Срезы промывали PBS между каждым этапом инкубации. Продукт иммунопероксидазной реакции визуализировали с помощью 3-амино-9-этилкарбазола (Aldrich) и фиксировали в формол-ацетате. Срезы контрастировали гематоксилином.
Электрофизиология.
Мыши находились на фоне гибридного штамма Blackswiss / 129. Большинство контрольных животных дикого типа были однопометниками мутантов. Учитывая, что LTD гиппокампа зависит от возраста, молодых мышей (возрастом 4 недели) использовали как в экспериментах по LTD, так и по депотенциации. Описан протокол электростимуляции и записи (23, 24). Во всех экспериментах использовался соответствующий возрасту контроль, и они проводились без учета генотипа. Поперечные срезы гиппокампа были быстро приготовлены и выдержаны в интерфейсной камере при 28 ° C, где они были подвергнуты субфузии с ACSF, состоящим из 124 мМ NaCl, 4,4 мМ KCl, 2,0 мМ CaCl 2 , 1,0 мМ MgSO 4 , 25 мМ NaHCO 3 , 1,0 мМ Na 2 HPO 4 и 10 мМ глюкозы. Интенсивность стимула регулировали для получения отклика амплитудой ≈1 мВ с начальным наклоном ≈ −0.5 мВ / мсек. Гомосинаптическая LTD вызывалась длительной низкочастотной стимуляцией (1 Гц в течение 15 мин). Для экспериментов с LTP каждые 50 секунд проверяли поле возбуждающего постсинаптического потенциала (ВПСП). ДП индуцировали тетанической стимуляцией (100 Гц дважды по 1 сек с интервалом 20 сек при интенсивности тестирования). Для экспериментов с посттетанической потенциацией (ПТП) ВПСП тестировали каждые 10 секунд до и после стимуляции.
Описан протокол электростимуляции и записи (23, 24). Во всех экспериментах использовался соответствующий возрасту контроль, и они проводились без учета генотипа. Поперечные срезы гиппокампа были быстро приготовлены и выдержаны в интерфейсной камере при 28 ° C, где они были подвергнуты субфузии с ACSF, состоящим из 124 мМ NaCl, 4,4 мМ KCl, 2,0 мМ CaCl 2 , 1,0 мМ MgSO 4 , 25 мМ NaHCO 3 , 1,0 мМ Na 2 HPO 4 и 10 мМ глюкозы. Интенсивность стимула регулировали для получения отклика амплитудой ≈1 мВ с начальным наклоном ≈ −0.5 мВ / мсек. Гомосинаптическая LTD вызывалась длительной низкочастотной стимуляцией (1 Гц в течение 15 мин). Для экспериментов с LTP каждые 50 секунд проверяли поле возбуждающего постсинаптического потенциала (ВПСП). ДП индуцировали тетанической стимуляцией (100 Гц дважды по 1 сек с интервалом 20 сек при интенсивности тестирования). Для экспериментов с посттетанической потенциацией (ПТП) ВПСП тестировали каждые 10 секунд до и после стимуляции. Измерялось облегчение реакции с помощью парных импульсов (PPF) с различными интервалами между импульсами (25–400 мс).Для экспериментов по депотенциации стимул для производства LTP составлял 100 Гц в течение 1 секунды, подаваемый дважды с интервалом в 20 секунд. За этим следовали низкочастотные стимулы 5 Гц в течение 3 мин для депотенцирования. В экспериментах с циклоспорином А (250 мкМ) срезы предварительно обрабатывали в течение 30 мин перед индукцией LTD.
Измерялось облегчение реакции с помощью парных импульсов (PPF) с различными интервалами между импульсами (25–400 мс).Для экспериментов по депотенциации стимул для производства LTP составлял 100 Гц в течение 1 секунды, подаваемый дважды с интервалом в 20 секунд. За этим следовали низкочастотные стимулы 5 Гц в течение 3 мин для депотенцирования. В экспериментах с циклоспорином А (250 мкМ) срезы предварительно обрабатывали в течение 30 мин перед индукцией LTD.
РЕЗУЛЬТАТЫ
Гистологический анализ кальциневрина Aα и Aβ.
Кальциневрин представляет собой гетеродимер, состоящий из каталитической (A; 61 кД) и регуляторной (B; 19 кД) субъединицы.Каталитическая субъединица существует в виде двух тесно связанных изоформ, Aα и Aβ, обе из которых экспрессируются в головном мозге. Кальциневрин Aα является преобладающей изоформой в гиппокампе, коре головного мозга, мозжечке и полосатом теле (1, 2, 16). Мышей с целевой мутацией в гене кальциневрина Aα получали стандартным методом гомологичной рекомбинации в эмбриональных стволовых клетках (16). Распределение кальциневрина Aα и Aβ в головном мозге мышей дикого типа и мутантных мышей исследовали иммуногистохимически с использованием антител к кальциневрину Aα и кальциневрину Aβ к пептидам (рис.1). В соответствии с предыдущими сообщениями (1, 2, 17) мы обнаружили, что у мышей дикого типа кальциневрин Aα высоко экспрессируется в областях CA1 и CA3 гиппокампа, а также в зубчатой извилине (рис. 1 A ). и C ). Белок кальциневрин Aα не был обнаружен в гиппокампе мутантных мышей (фиг. 1 B и D ). Кроме того, не было обнаружено заметной разницы в экспрессии кальциневрина Aβ между мышами дикого типа и мутантными мышами, что указывает на то, что кальциневрин Aβ не активируется во время развития у мутантных мышей.Эти данные свидетельствуют о том, что кальциневрин Aα не экспрессируется у мутантных мышей, а изоформа Aβ не компенсирует недостаток Aα у мутантных мышей. Как и ожидалось, активность кальциневрина (т.е. активность устойчивой к окадаовой кислоте и чувствительной к EGTA фосфатазы) была снижена в мозге мутантов более чем на 85% (18).
Распределение кальциневрина Aα и Aβ в головном мозге мышей дикого типа и мутантных мышей исследовали иммуногистохимически с использованием антител к кальциневрину Aα и кальциневрину Aβ к пептидам (рис.1). В соответствии с предыдущими сообщениями (1, 2, 17) мы обнаружили, что у мышей дикого типа кальциневрин Aα высоко экспрессируется в областях CA1 и CA3 гиппокампа, а также в зубчатой извилине (рис. 1 A ). и C ). Белок кальциневрин Aα не был обнаружен в гиппокампе мутантных мышей (фиг. 1 B и D ). Кроме того, не было обнаружено заметной разницы в экспрессии кальциневрина Aβ между мышами дикого типа и мутантными мышами, что указывает на то, что кальциневрин Aβ не активируется во время развития у мутантных мышей.Эти данные свидетельствуют о том, что кальциневрин Aα не экспрессируется у мутантных мышей, а изоформа Aβ не компенсирует недостаток Aα у мутантных мышей. Как и ожидалось, активность кальциневрина (т.е. активность устойчивой к окадаовой кислоте и чувствительной к EGTA фосфатазы) была снижена в мозге мутантов более чем на 85% (18).
Иммуногистохимическое определение кальциневрина Aα и кальциневрина Aβ в мозге мышей дикого типа и мутантных мышей с нокаутом кальциневрина Aα. A — D окрашены антисывороткой к пептиду кальциневрина Aα (R2929) у мышей дикого типа и мутантных мышей: кальциневрин Aα экспрессируется в гиппокампе мышей дикого типа ( A ) и, в частности, в CA1 области гиппокампа ( C ), но не у мутантных мышей ( B и D ).Кальциневрин Aβ [окрашенный антисывороткой к пептиду кальциневрина Aβ (R2948)] локализован в ядрах нейронов гиппокампа и в прилегающих трактах белого вещества. Интенсивность окрашивания сходна у мышей дикого типа [гиппокамп ( E ) и область CA1 ( G )] и мутантных мышей [гиппокамп ( F ) и область CA1 ( H )]. Коэффициент усиления составляет 11 ( A , B , E и F ) или 54 ( C , D , G и H ).
LTD является нормальным для мышей с дефицитом кальциневрина Aα.
Чтобы определить относительную роль кальциневрина Aα и Aβ в синаптической пластичности гиппокампа, мы изучили LTD, депотенцию и LTP в коллатеральном пути Шаффера у мышей с дефицитом кальциневрина Aα. Сначала мы оценили LTD, вызванную низкочастотной (1 Гц, 15 мин) стимуляцией (19). Предыдущие исследования показали, что фармакологические ингибиторы кальциневрина блокируют индукцию LTD (см.8, но см. Исх. 20). К нашему удивлению, LTD была нормальной у мышей с мутантом кальциневрина Aα. Стимуляция с частотой один герц в течение 15 мин вызвала у мутантных мышей такую же LTD синаптических ответов, что и у мышей дикого типа ( P > 0,05, различие незначительно; фиг. 2 a ). Поскольку сообщалось, что Ca 2+ / кальмодулин-зависимая протеинкиназа II регулирует частотно-зависимую функцию LTD гиппокампа (21), мы задались вопросом, может ли делеция кальциневрина Aα вызывать частотный сдвиг в ответ на электрическую стимуляцию. .Поэтому мы исследовали эффекты доставки одинакового количества импульсов (900) на разных частотах (5 и 10 Гц). На обеих частотах мыши дикого типа и мутантные мыши были неотличимы (рис. 2 c и d ). Эти результаты дополнительно подтверждают доказательства того, что кальциневрин Aα не является критическим для гомосинаптического LTD.
Рисунок 2LTD является нормальным для мышей с мутантом кальциневрина Aα и блокируется циклоспорином A. ( a1 ) Низкочастотная стимуляция (1 Гц, 15 мин) индуцировала LTD в области CA1 гиппокампа мышей дикого типа (белые квадраты; 61.6 ± 9,4%, n = 14 срезов / 10 мышей, t = 4,09, P <0,01 при сравнении потенциала через 25-30 минут после стимуляции со средним значением потенциала до стимуляции) или мутантных мышей (закрашенные квадраты ; 63,8 ± 12,0%, n = 9 срезов от 5 мышей, t = 3,02, P <0,05). ( a2 ) Репрезентативные записи EPSP до и через 30 минут после низкочастотной стимуляции у мышей дикого типа (верхний график) и мутантных (нижний график) мышей.( b ) LTD у мышей дикого типа и мутантных мышей блокировали циклоспорином A. ( b1 ) Предварительная обработка циклоспорином A блокировала LTD у мышей дикого типа (100,6 ± 4,0%, n = 6 срезов / 4 мыши ). LTD не была затронута в группах, получавших носитель (открытые квадраты, 64,6 ± 0,5%, n = 6 срезов / 4 мыши). ( b2 ) Предварительная обработка циклоспорином A блокировала LTD у мутантных мышей (закрашенные квадраты, 96,6 ± 5,1%, n = 6 срезов / 4 мыши). LTD не была затронута у мутантных мышей, получавших носитель (светлые квадраты, 64.5 ± 0,4%, n = 6 срезов / 5 мышей). ( c ) Низкочастотная стимуляция с частотой 5 Гц не влияла на синаптическую передачу. Ни LTP, ни LTD не индуцировались у дикого типа (пустые квадраты, 94,5 ± 14,1%, n = 6 срезов / 6 мышей) и мутантов (закрашенные квадраты, 103,9 ± 5,5%, n = 7 срезов / 7 мышей). мышей. ( d ) Стимуляция с частотой 10 Гц не влияла на синаптическую передачу. Стимуляция с частотой 10 герц не вызывала ни потенцирования, ни депрессии у дикого типа (светлые квадраты, 106.1 ± 9,7%, n = 11 срезов / 9 мышей) или мутантных (закрашенные квадраты, 98,1 ± 13,0%, n = 8 срезов / 6 мышей) мышей.
Депотенция блокируется у мышей с дефицитом кальциневрина Aα.
Участвует ли кальциневрин Aα в других формах синаптической депрессии? Исследования in vivo, и in vitro, демонстрируют, что депотенциация является формой синаптической депрессии, которая отличается от LTD (22). Поэтому мы исследовали депотенцию у мышей дикого типа и мутантных мышей кальциневрина Aα.У мышей дикого типа стимуляция тета-частоты (5 Гц в течение 3 минут), применяемая через 5 минут после индукции LTP, вызвала длительное подавление потенцированного синаптического ответа (фиг. 3 a ). Такая же стимуляция не оказывала значительного влияния на ответ в срезах, которые не подвергались LTP. Напротив, у мутантных мышей значительно снизилась депотенциация. Стимуляция с тета-частотой вызывала лишь кратковременную депрессию с последующим возвратом в состояние потенцирования (рис. 3 b ).Только тета-частотная стимуляция не вызывала потенцирования или депрессии (рис. 2 c ). Эти результаты демонстрируют, что кальциневрин Aα играет роль в гомосинаптической депотентации.
Рисунок 3Депотенция отменяется у мышей с мутантом кальциневрина Aα. ( a ) Стимуляция тета-частоты вызвала депотенцию у мышей дикого типа. Сильная тетаническая стимуляция (100 Гц, дважды по 1 сек с интервалом 20 сек) вызвала длительное усиление синаптического ответа (светлые квадраты).Пятигерцовая стимуляция (3 мин) вызвала длительную депрессию при доставке через 5 мин после индукции ДП (закрашенные квадраты, 91,4 ± 12,9%, n = 14 срезов / 10 мышей, t = 5,57, P <0,01 сравнение потенциала через 25-30 минут после столбняка с нормальным LTP без стимуляции 5 Гц). ( b ) Стимуляция с тета-частотой не вызывала депотенцию у мышей с мутантом кальциневрина Aα. Сильная тетаническая стимуляция вызвала такое же длительное усиление синаптических ответов (незакрашенные квадраты), как и у мышей дикого типа.Однако стимуляция с частотой 5 Гц вызвала лишь кратковременную депрессию, но полевой ВПСП вернулся на уровень потенцирования через 30 минут (закрашенные квадраты, 162,5 ± 14,1%, n = 13 срезов / 7 мышей, незначительно отличается по сравнению с LTP без Стимуляция 5 Гц на срезах мутантных мышей). ( c ) Стимуляция тета-частоты осуществлялась через 1 минуту после индукции LTP, вызванной депотенцией у мышей дикого типа (белые квадраты, 122,9 ± 15,1%, n = 5 срезов / 5 мышей), но не мутантных мышей (темные квадраты, 213.6 ± 20,5%, n = 7 срезов / 5 мышей, t = 3,28, P <0,005 при сравнении потенциала через 25-30 минут после стимуляции тета-частоты между мышами дикого типа и мутантными мышами). ( d ) Низкочастотная стимуляция (1 Гц, 15 мин) индуцировала аналогичные LTD у мутантов (закрашенные квадраты, 130,3 ± 6,3%, n = 6 срезов / 6 мышей) и дикого типа (светлые квадраты, 103,5 ± 23,3%, n = 6 срезов / 5 мышей, t = 1,45, незначительно отличается по сравнению с потенциалом через 25-30 минут после низкочастотной стимуляции между мышами дикого типа и мутантными мышами) при доставке через 5 минут после индукции LTP.
Чем можно объяснить снижение депотенцирования у мутантных мышей? Одна возможность состоит в том, что критический период обращения LTP становится короче вследствие мутации. Если бы это было так, то введение тета-частотной стимуляции ближе к концу столбняка могло бы быть более успешным в обращении вспять LTP. Чтобы проверить эту возможность, мы проводили стимуляцию тета-частоты через 1 мин после индукции LTP. Стимуляция тета-частоты снова не смогла обратить LTP у мутантных мышей ( n = 6; рис.3 c ), тогда как он реверсировал LTP у мышей дикого типа ( n = 5; фиг. 3 c ). Мы также проверили эффект стимуляции частотой 1 Гц (15 мин) через 1 мин после индукции LTP. Эта стимуляция индуцировала сходную LTD у мутантных мышей и мышей дикого типа даже после LTP (фиг. 3 d ), указывая на то, что участие кальциневрина Aα в депотентации зависит от частоты, но не от времени.
Фармакологическое ингибирование блоков кальциневрина, LTD.
Если кальциневрин Aα является критическим для депотенцирования, но не LTD, играет ли кальциневрин Aβ роль в LTD? Чтобы проверить эту идею, мы исследовали влияние ингибитора кальциневрина циклоспорина A на LTD у мышей с мутантом кальциневрина Aα.В соответствии с предыдущим сообщением (8) предварительная обработка циклоспорином А блокировала LTD у мышей дикого типа (рис. 2). Более того, циклоспорин А также отменял LTD у мутантных мышей (фиг. 2 b ). Одним из объяснений этого результата является то, что кальциневрин Aβ, в отличие от Aα, может быть избирательно важен для LTD гиппокампа. Однако настоящая работа не может полностью исключить возможность того, что фармакологические ингибиторы (например, FK506 и циклоспорин A) влияют на LTD, воздействуя на другие целевые белки, не связанные с кальциневрином (15, 25).
LTP и облегчение парных импульсов нормальны у мутантных мышей.
Фармакологические ингибиторы кальциневрина могут влиять на токи, опосредованные рецептором N -метил-d-аспартата (NMDA), и на индукцию LTP (12, 13, 26, 27). Чтобы изучить участие кальциневрина Aα в LTP, мы индуцировали LTP с помощью сильной тетанической стимуляции (две последовательности 100 Гц) как у мышей дикого типа, так и у мутантных мышей. Эта стимуляция вызывала быстрое и продолжительное усиление синаптического ответа, который оставался усиленным в течение по крайней мере 1 часа как у мышей дикого типа, так и у мутантных мышей ( P > 0.05) (Рис.4 a ). Мы не обнаружили явных аномалий в других аспектах синаптической передачи или простой краткосрочной формы синаптической пластичности. NMDA-опосредованные ВПСП, выделенные фармакологически с использованием CNQX, существенно не различались у мышей дикого типа ( n = 6) и мутантных мышей ( n = 7) ( P > 0,05, незначительно отличается по сравнению с двумя группами) . Более того, PTP, кратковременная потенциация и облегчение парных импульсов (PPF) были сходными у мутантных мышей и мышей дикого типа ( P > 0.05; Рис.4). Базальная синаптическая возбудимость также не пострадала. Залп EPSP, измеренный у мышей дикого типа ( n = 7) и мутантных мышей ( n = 7), был сходным ( P > 0,05). Наконец (рис. 4 c ), мы обнаружили, что LTD блокируется антагонистом рецептора NMDA AP-5 (100 мкМ), что указывает на то, что эта форма LTD зависит от активации рецептора NMDA.
Рисунок 4LTP, PTP и PPF не были затронуты у мышей с мутантом кальциневрина Aα. ( a ) Репрезентативные записи EPSP до и через 50-60 минут после сильной тетанической стимуляции у мышей дикого типа (верхний след) и мутантных (нижний след) мышей.( b ) LTP является нормальным у мышей с мутантом кальциневрина Aα. Сильный столбняк (100 Гц, две 1-секундные последовательности, доставленные с 20-секундными интервалами) индуцировал длительное усиление у дикого типа (открытые квадраты, 196,7 ± 12,4%, n = 9 срезов / 6 мышей, t = 7,80, P <0,001 при сравнении потенциала через 50-60 минут после столбняка со средним значением потенциала до столбняка) аналогичен таковому у мутантных мышей (заштрихованные квадраты, 203,4 ± 19,6%, n = 6 срезов / 5 мышей, t = 5,28, P <0.01). ( c ) PTP и STP индуцировали одиночный цикл тетанических стимулов (100 Гц, 1 сек) и сходны у мышей дикого типа и мутантных мышей. ПТР у мышей дикого типа (первое измерение после тетанической стимуляции; 162,1 ± 10,7%, n = 8 срезов / 4 мыши) и мутантных (темные квадраты, 172,5 ± 12,4%, n = 8 срезов / 5 мышей) были идентичный. ( d ) PPF у мышей дикого типа (незакрашенные квадраты) аналогичен PPF у мутантных мышей (закрашенные квадраты). В интервале 50 мсек, когда PPF максимален, PPF составлял 144.9 ± 2,8% (дикий тип, n = 11 срезов / 7 мышей) и 142,5 ± 3,9% (мутант, n = 17 срезов / 9 мышей). ( e ) Низкочастотная стимуляция (1 Гц, 15 мин) не вызывала LTD в области CA1 гиппокампа в присутствии 100 мкМ АР-5, антагониста рецептора NMDA ( n = 6, 114,2 ± 20,4% от контроля, без существенной разницы при сравнении потенциала через 25-30 минут после стимуляции со средним значением потенциала до стимуляции). ( Вставка ) Репрезентативные записи ВПСП до и через 30 минут после низкочастотной стимуляции в присутствии AP-5.
Поскольку было высказано предположение, что метаботропные рецепторы глутамата вносят вклад в депотенцию (28), мы также изучили функцию метаботропных рецепторов глутамата у мутантных мышей. Поэтому мы измерили эффект селективного агониста mGluR tACPD на базальную синаптическую передачу как у мутантных мышей, так и у мышей дикого типа. Одним из эффектов tACPD на синаптическую передачу является ингибирование потенциал-управляемых кальциевых каналов с помощью процесса, связанного с G-белком (29). Мы обнаружили, что tACPD (200 мкМ) оказывает аналогичное ингибирующее действие на синаптическую передачу у мутантных мышей ( n = 7; 49.0 ± 7,7% от контроля) и мышей дикого типа ( n = 4; 55,6 ± 7,3% от контроля) ( P > 0,05; незначительные различия при сравнении двух групп).
ОБСУЖДЕНИЕ
В настоящем исследовании мы объединили генетический и фармакологический подходы и предоставили убедительные доказательства того, что кальциневрин Aα необходим для генерации депотенциации в области CA1 гиппокампа. Эти данные подтверждают предыдущие сообщения о том, что механизм депотенцирования отличается от такового у LTD (9).Хотя для депотенцирования требуется кальциневрин Aα, наши фармакологические данные предполагают, что LTD может потребоваться кальциневрин Aβ. Однако мы не можем исключить потенциальное влияние фармакологических ингибиторов на другие целевые белки.
Недавние исследования показывают, что две различные формы LTD существуют в области CA1 гиппокампа: LTD, зависимая от рецептора NMDA, которая чувствительна к ингибированию протеинфосфатаз, и LTD, не зависящая от рецептора NMDA, которая устойчива к ингибиторам протеинфосфатазы (32 ).Если LTD, изучаемая в настоящем отчете, является независимой от рецептора NMDA формой LTD, можно было бы легко объяснить, почему LTD не затрагивается у мышей с дефицитом кальцинеруина Aα. Однако мы обнаружили, что LTD блокируется в присутствии 100 мкМ AP-5 (фиг. 4 и ). Следовательно, очевидно, что мы здесь имеем дело с опосредованной рецептором NMDA формой LTD.
Предыдущие фармакологические исследования с использованием селективных ингибиторов цАМФ-зависимой протеинкиназы A (PKA) и генетические исследования с использованием PKA RIβ- и Cβ1-дефицитных мышей (23, 24) продемонстрировали, что сигнальный путь цАМФ-PKA играет важную роль как в LTD, так и в депотенциации, но не в ранней фазе ДП (23, 24).Настоящие результаты предоставляют прямые доказательства того, что для депотенцирования избирательно требуется кальциневрин Aα, но не LTD. Таким образом, эти несколько генетических исследований показывают, что как LTD, так и депотенциация имеют общий отличительный молекулярный компонент. Необходимы дальнейшие исследования, чтобы выяснить, играют ли LTD и депотенциация разные роли в физиологических функциях центральной нервной системы.
Благодарности
Эта работа была частично поддержана грантами Национального института старения, Национального института психического здоровья и Медицинского института Говарда Хьюза.
Сноски
↵ * М.З. и W.Z. внес равный вклад в эту работу.
↵§ Нынешний адрес: Отделение анестезиологии, Медицинский факультет Вашингтонского университета, Сент-Луис, Миссури 63110.
↵‖ Нынешний адрес: Кафедра биохимии, Медицинский колледж Ханьянского университета, 17 Haengdang-dong, Sungdong-gu, Seoul 133-791, Korea.
†† Кому следует адресовать запросы на перепечатку.электронная почта: erk5 {at} columbia.edu.
СОКРАЩЕНИЯ
- LTP,
- долгосрочное потенцирование;
- LTD,
- длительная депрессия;
- ВПСП,
- возбуждающий постсинаптический потенциал;
- NMDA,
- N -метил-d-аспартат;
- PTP,
- посттетаническое потенцирование;
- PPF,
- облегчение парных импульсов;
- PKA,
- протеинкиназа A
- Принята 1 февраля 1999 г.
- Copyright © 1999, Национальная академия наук
Пористые нанокомпозитные мембраны на основе функционального ОГ с селективной функцией по адсорбции лития
Гибридные мембраны на основе функционального нанолиста GO для селективной адсорбции лития из соединения имели большой потенциал. Здесь была описана простая и эффективная методология для получения высокой стабильности, селективности и противообрастающих свойств. Что еще более важно, мы изучили селективную адсорбционную способность изготовленных мембран к литию в среде с высоким содержанием магния и низким содержанием лития.Результаты показали, что изготовленная мембрана демонстрирует отличную адсорбцию лития и высокую эффективность адсорбции. Изготовленные мембраны имели относительно более низкий угол смачивания (67,226 °) и более высокий поток через мембрану (13,62 мл · см −2 мин −1 ), что указывало на то, что изготовленная мембрана имела высокую гидрофильность, поскольку присутствие карбоксильных и гидроксильных групп ГО может усилить взаимодействие между мембраной и водой.Кроме того, были показаны результаты селективности по проницаемости (значения β были больше 7) и способности к регенерации (90,9% максимальной адсорбционной способности сохранялось после 10 циклов). Общие результаты позволяют предположить, что функциональный ГО может рассматриваться как эффективная добавка для неорганико-органических композитных мембран для селективной адсорбции лития. Стоит отметить, что эта методология может применяться для выборочной идентификации и адсорбции лития и является экологически чистой, поскольку не приводит к вторичному загрязнению.
У вас есть доступ к этой статье
Подождите, пока мы загрузим ваш контент… Что-то пошло не так. Попробуйте снова?Мануальный терапевт в Шарлотте, Северная Каролина — Greenapple Sports and Wellness, Шарлотта Мануальный терапевт :: Выборочная функциональная оценка движений (SFMA)
Выборочная функциональная оценка движений (SFMA)
— это серия из 7 тестов на движения всего тела, предназначенных для оценки основных моделей движений, таких как наклоны и приседания, у людей с известной скелетно-мышечной болью.Когда клиническая оценка начинается с точки зрения модели движений, у клинициста есть возможность выявить значимые нарушения, которые, казалось бы, не связаны с основной жалобой на опорно-двигательный аппарат, но способствуют связанной с ним инвалидности. Эта концепция, известная как региональная взаимозависимость, является отличительной чертой SFMA.
Оценка направляет клинициста к наиболее дисфункциональной, неболезненной модели движений, которая затем подвергается детальной оценке.Этот подход разработан в качестве дополнения к существующему экзамену и служит моделью для эффективной интеграции концепций осанки, мышечного баланса и основных моделей движений в опорно-двигательную практику. За счет устранения наиболее неблагополучных, неболезненных моделей, применение целевых вмешательств (мануальная терапия и лечебные упражнения) не подвергается неблагоприятному воздействию боли.
SFMA предлагает врачам новый подход к лечению боли и дисфункции. Наша стандартизированная клиническая модель обеспечивает изоляцию причины травмы и эффективное лечение.
Рассмотрение моделей движения
Нормальное движение достигается за счет интеграции основных моделей движений с адекватным балансом подвижности и устойчивости для удовлетворения требований выполняемой задачи.
Человеческая система будет мигрировать к предсказуемым моделям движений в ответ на боль или при наличии слабости, скованности или структурных аномалий. Со временем эти модели движений с ослаблением боли приводят к защитным движениям и страху перед движением, что приводит к клинически наблюдаемым нарушениям, таким как уменьшение ROM, изменение длины мышц и снижение силы.Изолированный или региональный подход к оценке или лечению не восстановит всю функцию. Функциональное восстановление требует практического знания функциональных паттернов и карты дисфункциональных паттернов, чтобы получить клиническую перспективу и разработать эффективную стратегию лечения.
Желательны безболезненные функциональные движения для участия в профессиональной деятельности и в повседневной жизни. Многие компоненты включают безболезненное функциональное движение, включая адекватную осанку, ROM, мышечную активность, моторный контроль и реакции равновесия.Нарушения каждого компонента могут потенциально изменить функциональное движение, что приведет к боли или как ее следствие. Используя SFMA, врач может определить ключевые функциональные модели движений и описать критические точки оценки, необходимые для эффективного восстановления функционального движения. Этот подход разработан, чтобы дополнить существующую модель обследования и вмешательства клинициста предписанием терапевтических упражнений на основе движений.
SFMA служит для эффективной интеграции концепций осанки, мышечного баланса и основных паттернов системы движений в опорно-двигательную практику.Кроме того, он обеспечивает обратную связь об эффективности программы лечебных упражнений, нацеленной на дисфункциональный паттерн движений и связанные с ними нарушения.
Выборочная оценка функциональных движений | Активная хиропрактика
Выборочная функциональная оценка движений (SFMA) — это серия из 7 тестов на движение всего тела, предназначенных для оценки основных моделей движений, таких как наклоны и приседания, у людей с известной скелетно-мышечной болью.Когда клиническая оценка начинается с точки зрения модели движений, клиницист имеет возможность идентифицировать значимые нарушения, которые, казалось бы, не связаны с основной жалобой на опорно-двигательный аппарат, но способствуют связанной с ним инвалидности. Эта концепция, известная как региональная взаимозависимость, является отличительной чертой выборочной функциональной оценки движений. Оценка направляет клинициста к наиболее дисфункциональной, неболезненной модели движений, которая затем подвергается детальной оценке.Этот подход разработан в качестве дополнения к существующему экзамену и служит моделью для эффективной интеграции концепций осанки, мышечного баланса и основных моделей движений в опорно-двигательную практику. Обращаясь к наиболее дисфункциональным неболевым образцам, на применение целевых вмешательств (мануальная терапия и лечебные упражнения) боль не оказывает неблагоприятного воздействия. Выборочная функциональная оценка движений предлагает врачам и физиотерапевтам новый подход к лечению боли и дисфункции.Наша стандартизированная клиническая модель обеспечивает изоляцию причины травмы и эффективное лечение.
Клиническое применение выборочной функциональной оценки движений
После того, как правила клинического прогнозирования и специфические для диагноза методы будут исчерпаны, применение модели выборочной функциональной оценки движений позволит индивидуально назначать упражнения и прогрессировать, что включает в себя концепцию региональной взаимозависимости. Как специалистам по опорно-двигательной системе важно, чтобы медицинские работники понимали взаимосвязь между сегментами тела и то, как нарушения в одной области тела могут отрицательно повлиять на функцию другой области тела.Подход к назначению лечебных упражнений с точки зрения модели движения позволяет логически и методично рассматривать концепции мышечного дисбаланса и региональной взаимозависимости. Смоделированный на основе выборочного тестирования тканей Cyriax, каждый тест SFMA оценивается как функциональный / безболезненный, дисфункциональный / безболезненный, функциональный / болезненный или дисфункциональный / болезненный. Цель клинициста — выявить наиболее дисфункциональный и неболезненный паттерн движений и разбить его, чтобы определить первопричину дисфункции.Это включает использование традиционных тестов на длину мышц и суставов, которые приводят к корректирующей мануальной терапии и упражнениям. Акцент делается на выявлении наиболее дисфункциональных паттернов. Модель призывает к тому, чтобы вмешательство было направлено только на безболезненные паттерны. Это гарантирует, что неблагоприятное воздействие боли на моторику не помешает корректирующим стратегиям.
Рассмотрение моделей движения
Нормальное движение достигается за счет интеграции основных моделей движений с адекватным балансом подвижности и устойчивости для удовлетворения требований выполняемой задачи.Человеческая система будет переходить к предсказуемым образцам движений в ответ на боль или при наличии слабости, скованности или структурных аномалий. Со временем эти модели движений с ослаблением боли приводят к защитным движениям и страху перед движением, что приводит к клинически наблюдаемым нарушениям, таким как уменьшение ROM, изменение длины мышц и снижение силы. Изолированный или региональный подход к оценке или лечению не восстановит всю функцию. Функциональное восстановление требует практического знания функциональных паттернов и карты дисфункциональных паттернов, чтобы получить клиническую перспективу и разработать эффективную стратегию лечения.Желательны безболезненные функциональные движения для участия в занятиях и образах жизни.
Многие компоненты включают безболезненное функциональное движение, включая правильную осанку, ROM, мышечную активность, моторный контроль и реакции равновесия. Нарушения каждого компонента могут потенциально изменить функциональное движение, что приведет к боли или как ее следствие. Используя выборочную оценку функциональных движений, клиницист может определить ключевые функциональные модели движений и описать критические точки оценки, необходимые для эффективного восстановления функционального движения.Этот подход разработан, чтобы дополнить существующую модель обследования и вмешательства клинициста предписанием лечебных упражнений на основе движений. Выборочная функциональная оценка движений служит для эффективной интеграции концепций осанки, мышечного баланса и основных паттернов двигательной системы в опорно-двигательную практику. Кроме того, он обеспечивает обратную связь об эффективности программы лечебных упражнений, нацеленной на дисфункциональные модели движений и связанные с ними нарушения.
Обратитесь к активному хиропрактику по телефону 804.560.9355, чтобы договориться о выборочной функциональной оценке движений.
.