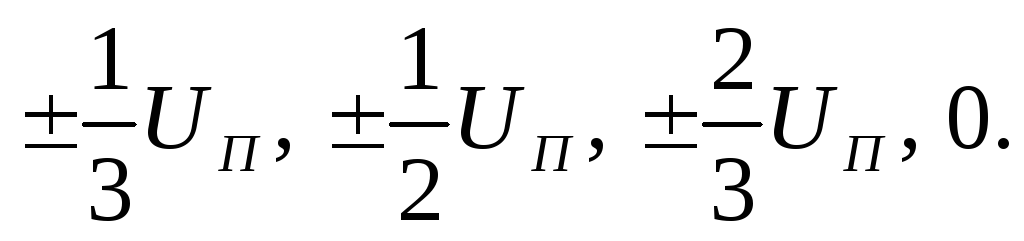Электрические явления в нервах и мышцах. Происхождение компонентов кривой однофазного тока действия
ГлавнаяРазноеПроисхождение компонентов кривой однофазного тока действияЭлектрические явления в нервах и мышцах
При определенных условиях в нервах и мышцах возникают электрические токи. Различают: 1) токи, или потенциалы, покоя и повреждения и 2) токи, или потенциал, действия – акционные точки.
Токи покоя
При отсутствии раздражения и, следовательно, возбуждения, существует разность потенциалов между наружной поверхностью цитоплазмы живой клетки (мембраной) и её внутренним содержимым.
Поверхностный слой цитоплазмы живой клетки, находиться в состоянии покоя, имеет положительный электрический заряд, а цитоплазма внутри клетки заряжена электроотрицательно. Поэтому, введя в клетку микроэлектрод и приложив другой электрод к поверхности клетки, можно отвести ток к регистрирующему прибору и измерить потенциал покоя, мембранный или клеточный потенциал.
Токи покоя, или клеточные потенциалы, обнаруживаются в неповрежденных живых клетках. Современными методами отведения электрического тока и его регистрации установлено, что токи покоя отдельных клеток животных, например гигантских нервных волокон кальмара, — 50-60 мв (милливольт), миелиновых нервных волокон амфибий – 70 мв, теплокровных – 90 мв, большинства тел нейронов – 80-100 мв, поперечнополосатых мышц амфибий – 80 мв, эпителия амфибий – 20 мв, волокон сердечной мышцы животных с постоянной температурой тела – 95 мв.
Токи повреждения
При повреждении живой ткани открывается доступ к внутреннему содержимому клеток, которое заряжено электроотрицательно. Поэтому если приложить электрод к неповрежденному участку ткани, находящемуся в состоянии покоя и, следовательно, заряженному электроположительно, а другой электрод приложить к поврежденному или перерожденному участку, заряженному электроотрицательно, то при соединении обоих электродов проводником тока с регистрирующим прибором можно измерить разность потенциалов.
Токи повреждения можно обнаружить на нервно-мышечном препарате. Если разрезать мышцу или любым способом повредить ее в одном участке (прижечь, раздавить и т. д.) и затем, приложив неполяризующиеся электроды, соединить срез мышцы или участок повреждения с целой поверхностью мышцы, не подвергающейся раздражению, то от целой поверхности к участку разреза или повреждения направится электрический ток. Сильно поврежденный участок всегда электроотрицателен по отношению к электроположительному неповрежденному участку. Следовательно, биоэлектрический ток повреждения, или альтерационный ток, возникает на границе целой и поврежденной частей ткани. Он обнаруживается и на некотором расстоянии от места повреждения, уменьшаясь с увеличением расстояния от места повреждения. Поэтому по месту происхождения его называют также демаркационным током. Однако ток повреждения появляется только тогда, когда отрицательно заряженный поврежденный участок соединяется проводником тока с положительно заряженным целым участком.
Величина тока повреждения меньше, чем клеточного потенциала, так как межклеточная и внеклеточная жидкости производят шунтирование (укорочение) при его измерении.
Разность потенциалов между целым и поврежденным участками скелетных мышц лягушки сразу же после повреждения равна 25-50 мв, наибольшая разность доходит до 80. В скелетных мышцах кролика токи повреждения равны 40 мв. В гладких мышцах токи повреждения меньше и быстрее уменьшаются до нуля. Разность потенциалов между целым и поврежденным участками седалищного нерва лягушки достигает 20-30 мв, седалищного нерва лошади — 6-16 мв, а седалищного нерва человека и обезьяны — 6-7 мв. Следовательно, токи повреждения нервов меньше, чем токи повреждения мышц. Особенно малы токи повреждения нервов человека и высших животных. Постепенно разность потенциалов уменьшается.
Когда нерв второго нервно-мышечного препарата набрасывается на первый нервно-мышечный препарат таким образом, чтобы он одновременно коснулся неповрежденного поврежденного участков мышцы, электрическая цепь замыкается. При этом мышца второго препарата сокращается в результате раздражения нерва током, возникающим в момент его замыкания между положительно заряженным нормальным участком мышцы первого препарата и отрицательно заряженным поврежденным ее участком.
Токи действия
Акционные токи, или токи (потенциалы) действия, возникают в раздражаемых тканях. Их появление характеризует возбуждение.
Для обнаружения тока действия нужно приложить один электрод к неповрежденному участку живой ткани, который находится в состоящих покоя и поэтому заряжен электроположительно, а другой — к раздражаемому участку и присоединить оба электрода к регистрирующему прибору. Раздражаемый участок при приложении к нему раздражителя немедленно или через несколько сотых миллисекунды становится электроотрицательным. Это падение потенциала’ доходит до максимума, а затем данный участок снова становится электроположительным. Различают однофазные и двухфазные токи действия.
Однофазный ток действия
При прикладывании одного из неполяризующихся электродов к неповрежденному участку нерва или мышцы, а другого — к поврежденному участку возникает электрический ток. Этот ток регистрируется при включении в цепь гальванометра или другого более чувствительною прибора.
Стрелка гальванометра отклонится в одну сторону вследствие электроотрицательного заряда поврежденного участка и электроположительного заряда нормального участка. Но есл
Физиология возбудимых тканей — Стр 2
В гиперкалиевом растворе потенциал действия существенно снижается. В гипернатриевом растворе его величина увеличивается. Для анализа потенциала действия используются также фармакологические вещества — они обладают способностью блокировать или калиевый или натриевый канал. При блоке натриевого канала потенциал действия снижается. Это очень важно в диагностике инфаркта миокарда, опухоли головного мозга и др. В зависимости от того, как электроды расположены к различным здоровым и больным участкам, можно зарегистрировать двухфазный или однофазный потенциал действия.
Двухфазный потенциал действия регистрируется в том случае, если электроды находятся на здоровом – неповрежденном участке ткани. Если к двум точка приложить отводящие электроды, а к другой точке (показано стрелкой) — раздражающие электроды, то при нанесении искусственного раздражения будет двухфазное колебание потенциала. Отводящие электроды соединены с регистрирующей аппаратурой. Механизм возникновения двухфазного потенциала действия заключается в том, что наружная поверхность клетки, мышечного или нервного волокна имеет положительный заряд, а цитоплазма – отрицательный.

Динамика электрических потенциалов в мышечном волокне
Поэтому при регистрации потенциала вначале будет просто прямая линия (а), б) волна возбуждения проходит через участок под первым электродом. Наружная поверхность мембраны в этом участке становится отрицательной и между электродами возникает разность потенциалов, стрелка отклоняется, кривая поднимается вверх, в) затем возбуждение занимает всю поверхность между электродами, разность потенциалов исчезает и стрелка возвращается обратно в начальное состояние, кривая идёт вниз. г) под первым электродом происходит реполяризация (положительный заряд), а под вторым электродом имеет место еще деполяризация и стрелка гальвонометра отклоняется уже в другую сторону, и кривая идёт вниз. д) возбуждение покидает пределы второго электрода, под ним происходит реполяризация, и стрелка гальванометра возвращается в начальное положение.
Это имеет важное значение для диагностики инфаркта миокарда, т.к. участок повреждения за ранее будет иметь отрицательный поверхностный заряд и волна возбуждения, которая будет распространяться на своем пути встретит измененный участок, а, следовательно, форма ЭКГ будет изменена.
Изменение возбудимости тканей при возбуждении.

местный Положительный следовой потенциал
процесс
Супернормальный период

Время, мс
местный процесс, фаза деполяризации, фаза реполяризацпи, отрицательный следовый потенциал, положительный следовый потенциал, а также фазы изменения возбудимости нервного волокна: фаза абсолютной рефрактерности, фаза относительной рефрактерности, супернормальная возбудимость, субнормальная возбудимость и исходный уровень возбудимости. В фазу абсолютной рефрактерности возбудимость падает до нуля. Это соответствует фазе деполяризации. Максимальная рефрактерность наблюдается в момент пика деполяризации. Если стойкая деполяризация будет вызвана каким-либо веществом, то ткань теряет возможность отвечать на приходящее следующее возбуждение. Практически, торможение может быть при стойкой гиперполяризации, при стойкой деполяризации и стойкой поляризации, когда под действием какой-либо причины поверхностный положительный заряд не меняется.
Периоду реполяризации соответствует фаза относительной рефрактерности. Здесь возбудимость постепенно восстанавливается. После относительной рефрактерности, наступает фаза супернормальной возбудимости — соответствует отрицательному следовому потенциалу, затем наступает субнормальная возбудимость — соответствует положительному следовому потенциалу, и затем возбудимость приходит к исходному уровню
Кривые одиночного сокращения (I) и изменения возбудимости (2) скелетной, сердечной и гладкой мышц.
: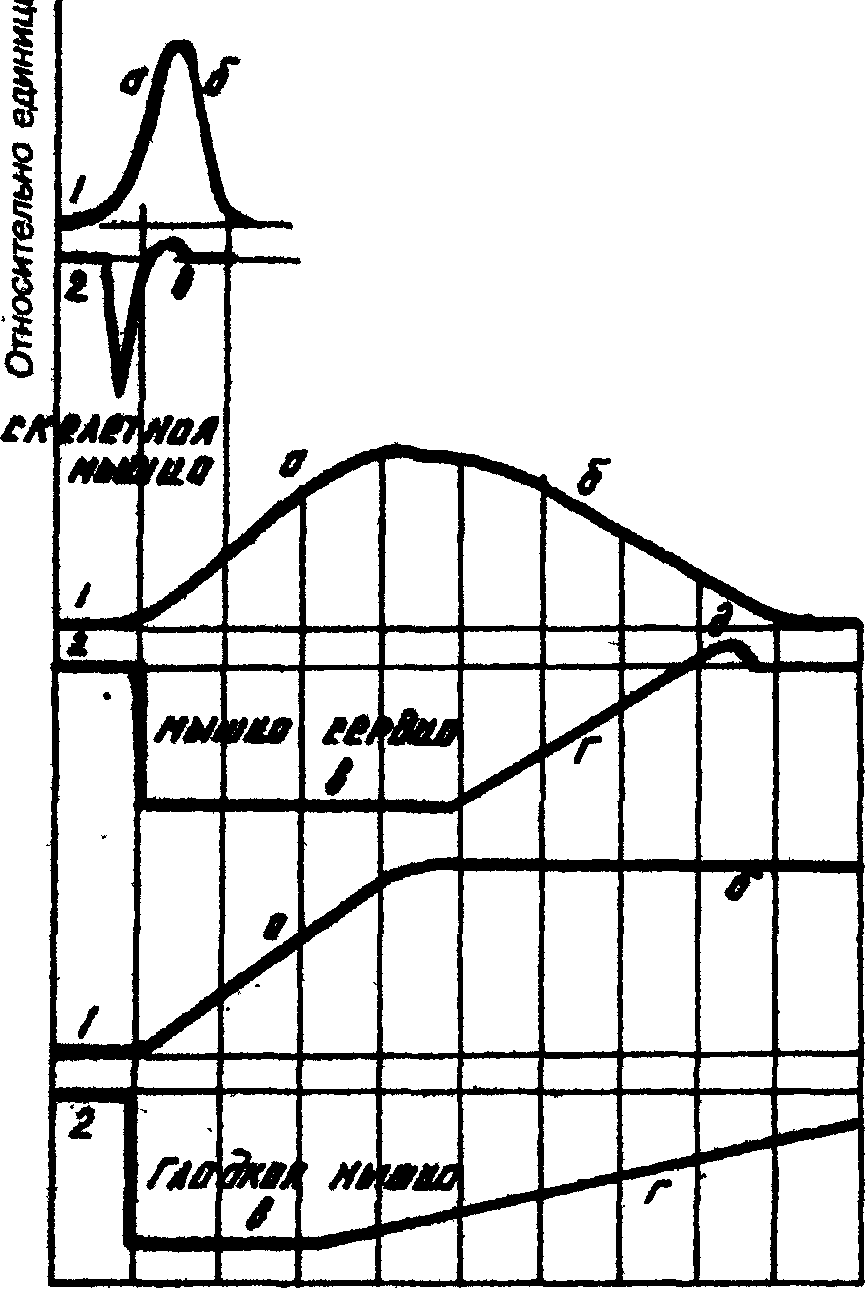
время, 0,1 с
а) период сокращения, б) период расслабления, в) фаза абсолютной рефрактерности, г) фаза относительной рефрактерности, д) фаза экзальтации (супернормальной возбудимости).
Разные мышцы обладают разной рефрактерностью и это свойство в значительной степени определяет особенности сократительной функции этих мышц.
Если взять постоянную частоту раздражающего тока, но постепенно увеличивать силу раздражения, то обнаружится, что с увеличением силы раздражения, ответная реакция будет увеличиваться. Такая же закономерность наблюдается в том случае, если увеличивать частоту наносимых раздражений при постоянной силе тока. Однако увеличение сокращения будет происходить до какой – оптимальной силы или частоты наносимых раздражений. Для оценки способности возбудимой ткани отвечать на раздражения разной частоты было введено понятие «лабильность» или функциональная подвижность (Н. Е. Введенский).
Под лабильностью понимают скорость протекания каждого цикла возбуждения или способность тканей воспроизводить без искажений частоту наносимых раздражений (для нервов лабильность ~ 1000 Гц, для скелетных мышц — ~250-500 Гц, для синапсов — ~ 100 Гц). Если частота наносимых раздражений больше лабильности, то не все импульсы будут воспроизводиться, а только те, которые не превышают величину лабильности (например, если на нерв нанести раздражения с частотой 2000 Гц, то ответов получим только 1000). При дальнейшем увеличении частоты, ответная реакция может исчезнуть. Для объяснения этого явления необходимо прибегнуть к понятиям абсолютной и относительной рефрактерности. Часть высокочастотных раздражений попадает на абсолютную рефрактерность, поэтому они ответа не вызывают. На основе лабильности Введенский разработал понятия об оптимуме и пессимуме силы и частоты раздражения. Та частота, при которой получается максимальный ответ — это оптимум частоты. Снижение ответной реакции, в связи с дальнейшим увеличением частоты наносимых раздражений называется пессимумом. Пессимум выражен тем больше, чем больше частота. Например, изменение возбудимости при одиночном сокращении скелетной мышцы:
При частоте больше 50 Гц возникает тетаничсское сокращение. Если наносить раздражение через интервалы одного цикла, то получится каждый раз одиночное сокращение. Если частоту увеличивать, интервалы между наносимыми раздражениями уменьшатся, и сокращение будет вначале ввиде зубчатого, а затем при дальнейшем увеличении частоты появится гладкий (сплошной) тетанус. Оптимум сокращения будет соответствовать фазе пика экзальтации — самой высокой возбудимости. При этом сила тока будет одна и таже, но так как возбудимость мышцы больше, то ответная реакция будет максимальной. При дальнейшем увеличении частоты, временные интервалы смешаются в фазу относительной рефрактерности, и часть импульсов при пессимуме попадает на эту фазу. Здесь возбуждение ниже, чем в фазу экзальтации, и ответная реакция будет ниже. Дальнейшее увеличение частоты приводит к попаданию импульсов на абсолютную рефрактерность. При этом нет ответа, т. к. в этот период полностью отсутствует возбудимость. Поэтому с лечебной целью используют токи высокой частоты, электроды прикладывают к коже, но мышцы не реагируют (не сокращаются), т. к. лабильность мышц намного ниже, чем частота тока УВЧ, и каждый стимул попадает на период абсолютной рефрактерности. Когда идёт множество импульсов из ЦНС по нерву к мышцам, то в зависимости от потребности в величине сокращения, к мышцам поступают импульсы различной частоты (например, чтобы поднять кусочек мела — поток нервных импульсов меньше, а чтобы поднять стул — больше, при этом сокращаются больше миофибрилл, и ответная реакция возрастает). Введенский, базируясь на учениях о лабильности, пессимуме и оптимуме, открыл явление парабиоза. Он брал нервно-мышечный препарат и раздражал нерв с использованием различной силы тока, и записывал сокращение мышцы. Ответная реакция при этом полностью укладывалась в «закон силы», т. е. с увеличением силы раздражения ответная реакция усиливалась. После этого он накладывал на нерв ватку, смоченную кокаином, и снова раздражал нерв. Он выявил, что обнаруживается фазное изменение возбудимости и проводимости: I) уравнительная фаза: здесь происходит выравнивание всех ответов, на все виды раздражений — одинаковый ответ, 2) парадоксальная фаза — слабое раздражение даёт больший ответ, чем раздражители средней и большой силы, 3) тормозящая стадия — нет ответной реакции ни на одно раздражение. Это происходит потому, что в участке альтерации (где ватка с-кокаином) постепенно снижается лабильность. Это приводит: I) в уравнительную фазу — к пропусканию определённого числа импульсов, а избыточное число (которое больше лабильности альтерированного участка) блокируется и до мышцы доходит одинаковое число импульсов, 2) в парадоксальную фазу происходит дальнейшее снижение лабильности, и ответ извращённый: малое число импульсов проходит, но с увеличением силы и частоты раздражения, часть импульсов блокируется, ответная реакция снижается. Дальнейшее увеличение силы и частоты приводит к большей блокаде импульсов — происходит по принципу пессимума. 3) в тормозящую фазу лабильность падает дальше и проводимость в участке альтерации вообще прекращается, и импульсы не доходят до мышцы. На этом основана новокаиновая анестезия. Действие новокаина базируется на том, что лабильность рецепторов и афферентных проводников снижается. Импульсы не доходят до центра и боль не чувствуется.
Введенский впервые обосновал теорию единства возбуждения и торможения. Он рассматривал торможение как частный случай возбуждения, но особого — не распространяющегося, стационарного. По Введенскому, есть возбуждение импульсное (обычное), а под влиянием альтерирующего агента возникает местное локальное нераспростравяющееся возбуждение. Стадии парабиоза являются результатом взаимодействия двух возбуждений – импульсного и локального (стационарного). Развитие парабиоза и возникновение торможения следует рассматривать как вторичное торможение, обусловленное взаимодействием двух возбуждений. Явление парабиоза имеет универсальный характер, и он может развиться в разных отделах ЦНС и в периферических нервах при действии экстремальных факторов, больших доз лекарственных веществ. Парабиотические явления (в виде фазовых явлений) могут быть и в высшей нервной деятельности.
Физиологические свойства нервных волокон. По скорости проведения возбуждения нервные волокна подразделяются на несколько групп. Наибольшее признание получила классификация нервных волокон по Эрлангеру-Гассеру. Согласно этой классификации выделены 3 основные группы нервных волокон – А, В, С. В свою очередь группа А делится на несколько подгрупп ( – альфа, – бета, g – гамма и — дельта). Наибольшая скорость проведения возбуждения (70-120 м /сек) в группе А альфа – такую скорость имеют первичные афференты мышечных веретен, двигательные волокна скелетных мышц. Группа А — — это кожные афференты прикосновения и давления имеют скорость проведения возбуждения 30-70 м/сек. Группа A -гамма имеет скорость 15-30 м/сек – это двигательные волокна мышечных веретен. Группа А –дельта имеет скорость проведения возбуждения 12 – 30 м/сек, такую скорость имеют кожные афференты температуры и боли (первичная боль). Группа В имеет скорость 3 – 15 м/сек. это главным образом симпатические преганглионарные волокна. Группа С имеет скорость 0,5 – 2 м/сек. – это кожные афференты боли (вторичная, медленная боль) и симпатические постганглионарные волокна (немиелинизированные).
Аксоток. Нервные волокна имеют своеобразное строение — микротрубочки, по которым перемещаются вещества от нервной клетки на периферию (антероградный поток) и от периферии к центру (ретроградный аксоток). Различают быстрый (около 410 мм в сутки) и медленный (примерно в 2 раза медленнее) аксоток. За счёт него от центра к периферии распространяются биологически активные вещества. Аксон, диаметр которого составляет всего несколько микронов, может достигать длины одного метра и более, и движение белков путем диффузии от. ядра к дистальному концу аксона заняло бы годы. Давно известно, что когда какой-либо из участков аксона подвергается констрикции, часть аксона, расположенная проксимальнее, расширяется. Это выглядит так, как будто в аксоне блокирован центробежный поток. Такой поток — быстрый аксонный транспорт может быть продемонстрирован движением радиоактивных маркеров в эксперименте. Лейцин, меченный радиоактивной меткой, инъецировали в ганглий дорсального корешка, и затем со 2-го по 10-й час измеряли радиоактивность в седалищном нерве на расстоянии 166 мм от тел нейронов. За 10 часов пик радиоактивности в месте инъекции менялся незначительно. Но волна радиоактивности распространялась по аксону с постоянной скоростью около 34 мм за 2 ч, или 410 мм • сутки. Показано, что во всех нейронах гомойотермных животных быстрый аксонный транспорт осуществляется с такой же скоростью, причем ощутимых различий между тонкими, безмиелиновыми волокнами и наиболее толстыми аксонами, а также между моторными и сенсорными волокнами не наблюдается. Тип радиоактивного маркера также не влияет на скорость быстрого аксонного транспорта; маркерами могут служить разнообразные радиоактивные вещества.
Если проанализировать периферическую часть нерва, чтобы понять природу переносчиков радиоактивности, то такие переносчики обнаруживаются главным образом во фракции белка, а также в составе медиаторов и свободных аминокислот. Описанный выше быстрый аксонный транспорт является антероградным, т. е. направленным от тела клетки. Показано, что некоторые вещества движутся с периферии к телу клетки с помощью ретроградного транспорта. Например, ацетилхолинэстераза транспортируется в этом направлении со скоростью 2 раза меньшей, чем скорость быстрого аксонного транспорта. Маркер, часто используемый в нейроанатомии пероксидаза хрена — также перемещается ретроградным транспортом. Ретроградный транспорт, вероятно, играет важную роль в регуляции белкового синтеза в теле клетки. Через несколько дней после перерезки аксона в теле клетки наблюдается хроматолиз, что свидетельствует о нарушении белкового синтеза. Время, требующееся для хроматолиза, коррелирует с длительностью ретроградного транспорта от места перерезки аксона до тела клетки. За счёт антероградного тока происходит дифференциация тканей (например, мышц). Это имеет большое биологическое значение. Есть мышцы фазные (мышцы конечностей) и тонические (поддерживают позу). В эксперименте установлено, что если перерезать нервы, иннервирующие эти мышцы, а затем произвести перекрест иннервации, то есть центральный конец нерва, иннериврующего фазные мыщы, подшить к нерву, иннервирующему тонические мышцы, то после проростания нервов, фазные мышцы начинают функционировать как тонические, а тонические – как фазные. Меняется их структура, так как за счёт аксотока обеспечивается трофическая функция двигательных нервов. За счет ретроградного тока нейротропные вещества поступают от периферии к центру, оказывая трофическое влияние на саму нервную клетку. Путем ретроградного транспорта могут поступать к телу нервной клетки токсины, а также некоторые химические вещества, используемые в производственных условиях. Быстрый аксонный транспорт требует значительной концентрации АТФ. Такие яды, как колхицин, разрушающий микротрубочки, также блокируют быстрый аксонный транспорт. Из этого следует, что в рассматриваемом нами транспортном процессе везикулы и органеллы движутся вдоль микротрубочек и актиновых филаментов; это движение обеспечивается малыми агрегатами молекул динеина и миозина, действующих с использованием энергии АТФ.
Быстрый аксонный транспорт может участвовать и в патологических процессах. Некоторые нейротропные вирусы (например, вирусы герпеса или полиомиелита) проникают в аксон на периферии и движутся с помощью ретроградного транспорта к телу нейрона, где размножаются и оказывают свое токсическое действие. Токсин столбняка — белок, который продуцируется бактериями, попадающими в организм при повреждениях кожи, захватывается нервными окончаниями и транспортируется к телу нейрона, где он вызывает характерные мышечные спазмы. Известны случаи токсического воздействия на сам аксонный транспорт, например воздействие промышленным растворителем акриламидом. Кроме того, полагают, что патогенез авитаминоза «бери-бери» и алкогольной полинейропатии включает нарушение быстрого аксонного транспорта.
Как уже говорилось выше, помимо быстрого аксонного транспорта в клетке, существует и довольно интенсивный медленный аксонный транспорт. Тубулин движется по аксону со скоростью около 1 мм/сут, а актин быстрее — до З мм/сут. С этими компонентами цитоскелета мигрируют и другие белки; например, ферменты, по-видимому, связаны с актином или тубулином. Скорости перемещения тубулина и актина примерно согласуются со скоростью роста, обнаруженной для механизма, когда молекулы включаются в активный конец микротрубочки или микрофиламента. Следовательно, этот механизм может лежать в основе медленного аксонного транспорта. Скорость медленного аксонного транспорта примерно соответствует также скорости роста аксона, что, по-видимому, указывает на ограничения, накладываемые структурой цитоскелета на второй процесс.
Следует подчеркнуть, что клетки ни в коем случае не являются статичными структурами, каковыми они кажутся, например, на электронно-микроскопических фотографиях. Плазматическая мембрана и особенно органеллы находятся в постоянном быстром движении и постоянной перестройке; только поэтому они способны функционировать. Далее, это не простые камеры, в которых протекают химические реакции, а высокоорганизованные конгломераты мембран и волокон, в которых реакции протекают в оптимально организованной последовательности.
Физиологические свойства нервных волокон:.1) возбудимость — способность реагировать на приходящий импульс. 2) проводимость — способность распространять импульсы от одного участка к другим. Эти свойства зависят от структуры нервного волокна. Все нервные волокна подразделяются на I) мякотные — имеют миелиновую оболочку, перехваты Ранвье, имеющие важное значение для передачи возбуждения. Сама миелиновая оболочка является мощным биологическим изолятором. Через неё возбуждение не перескакивает с одного нервного волокна на соседние. Поэтому проходящий импульс неэффективен для соседних волокон. 2) безмиелпновые нервные волокна — передача возбуждения в них происходит по поверхности нерва через изменение поверхностного заряда. Обычно нервный ствол содержит большое количество нервных волокон. Безмиелиновые волокна в нём находятся среди миелиновых. Законы проведения возбуждения по нервному волокну: I) закон физиологической целостности. Анатомическая целостность — когда на нервном волокне сохранены все структуры. Функциональная целостность может быть нарушена действием каких-либо факторов без повреждения струкутуры, например, парабиоз. Чтобы возбуждение прошло по волокну, должна быть его физиологическая целостность. 2) закон двухстороннего проведения возбуждения: если на нерв поставить два гальванометра и наносить раздражение между приборами, то возникший потенциал действия регистрируется как справа, так и слева. В любом целостном организме возбуждение фактически идёт в одном направлении (от аф-ферентного канала через центр к эфферентному, т. к. синапсы проводят возбуждение односторонне). 3) закон бездекрементного проведения возбуждения (без ослабления) — вне зависимости от размеров и длины нейрона, возбуждение не теряет своей силы — в разных участках величина потенциала действия будет одинакова. 4) закон изолированного проведения возбуждения — по каждому нервному волокну возбуждение передаётся только вдоль этого волокна. Поперечная передача с одного нервного волокна на другое не происходит. Без этого не было бы координации движений. Нарушение этого правила бывает при травме нерва и его сопоставлспии. 5) закон скачкообразного проведения возбуждения (сальтаторного) – такая передача возбуждения происходит только в миэлиновых нервных волокнах. Большое значение имеют перехваты Ранвье, т. к. возбуждение перескакивает с одного на другой перехват или даже минуя один перехват. Поэтому такие нервные волокна имеют наибольшую скорость проведения возбуждения. 6) безмиэлиновые волокна проводят возбуждение по поверхности. Возбужденный участок заряжается электроотрицательно (деполяризация) и эта волна распространяется вдоль нервного волокна.
. Механизм передачи возбуждения с нерва на мышцу.
Возбуждение, возникающее в ЦНС, через эфферентные каналы доходит до скелетных мышц. Есть 2 механизма передачи возбуждения с нерва на рабочий орган: I) химический – в результате выработки медиаторов 2) электрический, когда потенциал действия с пресинаптической мембраны перескакивает на постсинаптическую мембрану и вызывает ее деполяризацию. Основным условием является расстояние между пре- и постсинаптическими мембранами: если оно больше 0,2 нм, то передача будет химическая, а если меньше, то потенциал с пресинаптической мембраны переходит на постсинаптическую, происходит ее деполяризация и мышца сокращается. Эта передача мало распространена. Мионевральные синапсы передают возбуждение на скелетные и гладкие мышцы, за счет синапсов происходит передача возбуждения в ЦНС, включая кору. Медиаторы могут быть различные. Самый распространённый: а) ацетилхолин — для скелетных мышц, в парасимпатических нервах, холинэргические синапсы ЦНС; б) адренергические синапсы на периферию передают возбуждение за счет норадреналина, который, выделяясь на окончаниях симпатических нервов, влияет на сердце, сосуды, желудочно-кишечный тракт. В симпатических ганглиях передача возбуждения с преганглионарных на постганглионарные нейроны происходит за счёт ацетилхолина. Адренергические синапсы распространены в ЦНС (особенно в стволе мозга). Симпатическая иннервация головного мозга происходит главным образом за счёт синего пятна, которое синтезирует адренергические вещества; в) серотонинергические структуры.- вырабатывают серотонин (в основном ядра шва мозга), г) специфические синапсы — чувствительны к определённым нейропептидам, синтезируемым в структурах головного мозга, в слизистой ЖКТ, и в надпочечниках. Они действуют на периферические и центральные структуры.
Механизм химической передачи:
Строение синапса: I) терминаль — окончание двигательного нерва, 2) бляшка, 3) пузырьки, содержащие медиатор, 4) пресинаптическая мембрана, 5) синаптическая щель, 6) постсинаптическая мембрана. При распространении импульса по нейрону возбуждение по термнали доходит до синаптической бляшки, в результате чего из пузырьков через пресинаптическую мембрану в синаптическую щель поступает медиатор. На постсинаптической мембране имеются специальные холинорецепторы (для медиатора ацетилхолина) или адренорецепторы (для норадреналина). На окончаниях двигательных нервов выделяется медиатор ацетилхолин, когда он поступает в синаптическую щель вызывает повышение проницаемости постсинаптической мембраны для ионов Nа+. Возникновение потока ионов Nа+ через постсинаптическую мембрану вызывает деполяризацию ее и образуется возбуждающий постсинаптический потенциал (ВПСП) или потенциал концевой пластинки (ПКП). На пресинаптической мембране имеются активные зоны, то есть участки пресинаптической мембраны, где больше всего происходит выброс ацетилхолина в синаптическую щель. Рядом расположены неактивные участки. Виды секреции ацетилхолина: I) квантовая секреция (или вызванная возбуждением) — с помощью импульса — является основным фактором, вызывающим деполяризацию постсинаптической мембраны и сокращение мышц. 2) спонтанная секреция — когда поступление импульса не обязательно. Это приводит к возникновению локального
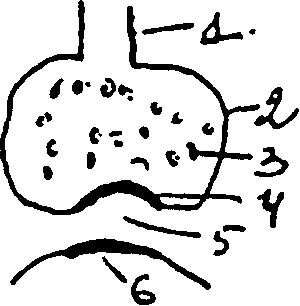
потенциала, который незначительный и не вызывает сокращения мышцы. Он регистрируется в виде небольшой разности потенциалов, в) неквантовая секреция — на неё приходится около 30% выделившегося медиатора, но она не сопровождается деполяризацией постсинаптической мембраны, и не вызывает сокращение мышцы. За счёт неё обеспечивается трофика. Регуляция: I) пресинаптическая ауторегуляция секреции ацетилхолина — в бляшке терминали имеются механизмы, способные регулировать выброс ацетилхолина. Они могут оказывать двоякое действие на функциональное состояние синапса: I) десенситизация — снижение чувствительности синаптических рецепторов. Когда чувствительность их снижена до состояния невосприимчивости возбуждения, развивается состояние синаптического торможения. Оно обусловлено снижением чувствительности холинорецепторов, расположенных на постсинаптической мембране. Может быть пресинаптическое торможение, которое обусловлено снижением выработки и выброса медиатора или блокадой их. Поэтому в фармакологии есть блокаторы, угнетающие пресинаптические структуры, или снижающие чувствительность постсинаптических мембран. 2) синаптическая потенциация — повышение возбудимости. Спонтанная секреция ацетилхолина может привести к этим явлениям: происходит повышение возбудимости холинорецепторов, и при выделении кванта медиатора, он действует на состояние повышенной возбудимости и вызывает большую ответную реакцию. Это происходит в зависимости от выполнения человеком работы. Медиатор ацетилхолин, поступивший в синаптическую щель, подвергается быстрому разрушению ферментом ацетилхолинэстеразой. Если бы его не было. то ацетилхолин долго бы раздражал рецепторы постсинаптических мембран, и не было бы адекватной координации действия мышцы. Поэтому новое сокращение происходит только при поступлении нового кванта медиатора. Значительная часть холина, образующегося при разрушении ацетилхолина, принимает участие в ресинтезе новых квантов ацетилхолина. Новые порции медиатора расположены дальше от пресинаптической мембраны, а ближе расположены зрелые пузырьки с ацетилхолином, которые потом освобождают медиатор. В основе возникновения потенциала на постсинаптической мембране лежит ионно-мембранный механизм:
Введение в физиологию. Основные характеристики живого организма.
План
практических занятий по нормальной физиологии для студентов 1
курса стоматологического факультета на весенний семестр 2011 – 2012 учебного года
Занятие № 1.
Введение в физиологию. Основные характеристики живого организма.
(10 февраля 2012 г.)
Занятие № 2.
Электрические явления в нервно-мышечной системе.
(17 февраля 2012 г.)
Занятие № 3.
Физиология нервов и нервно-мышечного синапса.
(24 февраля 2012 г.)
Занятие № 4.
Физиология мышц. Механизм мышечного сокращения.
(2 марта 2012 г.)
Занятие № 5.
Сердечный цикл и его фазы. Автоматия сердца.
(9 марта 2012 г.)
Занятие № 6.
ФИЗИОЛОГИЧЕСКИЕ СВОЙСТВА СЕРДЕЧНОЙ МЫШЦЫ.
(16 марта 2012 г.)
Занятие № 7.
Физические проявления деятельности сердца. Физиологические основы электрокардиографии.
(23 марта 2012 г.)
Занятие № 8.
ДВИЖЕНИЕ КРОВИ ПО СОСУДАМ. КРОВЯНОЕ ДАВЛЕНИЕ.
(30 марта 2012 г.)
Занятие № 9.
Нервно — гуморальная регуляция сердца и сосудов.
(6 апреля 2012 г.)
Занятие № 10.
Рефлекторная регуляция сердечно — сосудистой системы.
(13 апреля 2012 г.)
Занятие № 11.
(20 апреля 2012г.)
Итоговое занятие по теме: Сердечно — сосудистая система.
Занятие № 12.
Общая физиология желез внутренней секреции.
(27 апреля 2012 г.)
Занятие № 13.
Частная физиология желез внутренней секреции.
(4 мая 2012 г.)
Занятие № 14.
Челюстно-лицевая область. Часть первая.
(11 мая 2012 г.)
Занятие № 15.
Челюстно-лицевая область. Часть вторая.
(18 мая 2012 г.)
Занятие № 16.
Общая физиология сенсорных систем. Слуховой анализатор. Кожная и болевая рецепция.
(25 мая 2012г.)
Занятие № 17.
Зрительный анализатор. Вкусовой и обонятельный анализаторы.
(1 июня 2012 г.)
Занятие № 18.
Физиология больших полушарий. Понятие о высшей нервной деятельности (ВНД).
(8 июня 2012 г.)
Занятие № 19.
Учение о высшей нервной деятельности. Физиологические основы памяти, сна и эмоций. Понятие о функциональных системах.
(15 июня 2012 г.)
Занятие № 1.
Введение в физиологию. Основные характеристики живого организма.
(10 февраля 2012 г.)
1. Предмет и метод физиологии.
2. Связь физиологии с другими дисциплинами, ее значение для медицины.
3. Основные физиологические функции и процессы, протекающие в живом организме (обмен веществ и энергии, рост, размножение, пищеварение, дыхание, кровообращение, работа нервной систнемы). Понятие о гомеостазе.
4. Показатели, характеризующие нормальное состояние организма: температура тела, частота и характер пульса, частота дыхания, артериальное давление, осмотическое давление крови, Рн и др.)
5. Понятие о механизмах связи и регуляции функций в организме.
6. Общие свойства возбудимых тканей: раздражимость и возбудимость.
7. Виды раздражителей.
8. Порог раздражения (реобаза). Полезное время действия тока. Хронаксия. Кривая зависимости силы и времени действия тока.
9. Изменение возбудимости при возбуждении: рефрактерная фаза (абсолютная и относительная), фаза повышенной (супернормальной ) и фаза пониженной (субнормальной) возбудимости.
10. Гальванические явления, возникающие при наличии разнородных металлов в полости рта, значение для стоматологии.
Занятие № 2.
Электрические явления в нервно-мышечной системе.
(17 февраля 2012 г.)
1. Современные представления о строении и функции мембран клеток.
2. Ионные каналы клеточных мембран, классификация.
3. Электрические явления в возбудимых тканях.
4. Токи покоя (мембранный потенциал, ток повреждения, метаболический ток).
5. Происхождение мембранного потенциала (значение ионной асимметрии во внеклеточной и внутриклеточной жидкости). Ионные градиенты клетки, их механизмы. Уравнение Нернста.
6. Натриево-калиевый насос и его роль в поддержании градиента ионных концентраций в клетке в состоянии покоя.
7. Современные представления о процессе возбуждения. Локальный ответ. Критический уровень деполяризации при действии раздражителей. Понятие о поляризации мембраны, ее деполяризации (реверсии заряда), реполяризации и гиперполяризации.
8. Потенциал действия и его фазы. Условия регистрации двухфазного и однофазного тока действия.
9. Происхождение компонентов кривой однофазного тока действия, их продолжительность. Мембранно-ионная теория происхождения тока действия.
10. Соотношение фаз возбудимости с фазами потенциала действия.
11. Работа натриево-калиевого насоса после возбуждения.
Занятие № 3.
Физиология нервов и нервно-мышечного синапса.
(24 февраля 2012 г.)
1. Значение нервной системы в механизмах регуляции функций в организме. Функция нервов.
2. Строение периферических нервных волокон.
3. Физиологические свойства нервных волокон (возбудимость и проводимость).
4. Механизм проведения возбуждения в мякотных и безмякотных нервных волокнах.
5. Классификация нервных волокон по их диаметру, скорости проведения возбуждения и особенностям потенциала действия ( группы А, В и С).
6. Законы проведения возбуждения в нервах ( закон анатомической и физиологической целостности нервного волокна, закон двухстороннего проведения возбуждения и закон изолированного проведения возбуждения ).
7. Нервно-мышечный синапс, структура и свойства, медиатор. Понятие о холино — рецепторе.
8. Механизм передачи возбуждения с нерва на мышцу, Потенциал концевой пластинки (ПКП) и его свойства. Значение фермента холинэстеразы.
9. Возникновение потенциала действия в мышечном волокне.
10. Понятие о лабильности (Н.Е. Введенский).
11. Парабиоз (Н.Е.Введенский). Сущность и значение учения о парабиозе для стоматологии. Физиологические основы проводникового обезболивания.
12. Физиологические основы местного обезболивания.
Занятие № 4.
ФИЗИОЛОГИЯ МЫШЦ. МЕХАНИЗМ МЫШЕЧНОГО СОКРАЩЕНИЯ.
(2 марта 2012 г.)
1. Физиологическая роль мышечной системы. Классификация мышц.
2. Морфо-функциональные особенности исчерченных мышц.
3. Ультраструктура миофибрилл: тонкие и толстые протофибриллы, их взаимное расположение в саркомере.
4. Сократительные белки (миозин и актин) и регуляторные белки (тропомиозин и тропонин) в составе протофибрилл, их свойства.
5. Понятие о саркоплазматическом ретикулюме (СПР). L- система продольных трубочек, цистерны СПР, их значение как депо ионов кальция. Т- система поперечных трубочек СПР, мембраны миофибрилл, взаимодействие с L — системой.
6. Современная теория мышечного сокращения и расслабления.
7. Биоэлектрические, химические и тепловые процессы в мышцах.
8. Электромеханическая связь возбуждения и сокращения мышцы.
9. Физические и физиологические свойства мышц.
10. Режимы и виды сокращений мышц (одиночные и тетанические).
11. Одиночное сокращение и его фазы. Изменение возбудимости при возбуждении мышцы.
12. Явление суммации сокращений.
13. Тетанические сокращения: зубчатый и гладкий тетанус.
14. Понятие о лабильности (Н.Е. Введенский). Оптимум и пессимум силы и частоты для сокращения мышцы.
15. Основные отличия в строении и функции скелетных и гладких мышц внутренних органов.
16. Методы изучения возбудимости мышц и нервов в стоматологии. Электроодонтометрия, её значение в стоматологии.
17. Гнатодинамометрия. Закон средних нагрузок, особенности жевательных и мимических мышц. Контрактура жевательных и мимических мышц и её последствия. Электромиография, её значение для стоматологии.
Занятие № 5.
Сердечный цикл и его фазы. Автоматия сердца.
(9 марта 2012 г.)
1. Значение кровообращения для организма.
2. Развитие учения о кровообращении. Схема кровообращения взрослого человека.
3. Строение сердца, его гемодинамическая функция. Клапанный аппарат сердца, его значение.
4. Частота сокращений сердца. Понятие о сердечном цикле, его продолжительность, основные фазы: систола предсердий, систола желудочков и общая пауза.
5. Фазовый анализ сердечного цикла.
6. Изменения давления и объема крови в полостях сердца во время фаз сердечного цикла.
7. Проводящая система сердца, ее морфологические особенности. Узлы: синусный узел, атриовентрикулярный узел; внутрипредсердные тракты, пучок Гиса, его ножки, волокна Пуркинье.
8. Понятие об автоматии. Современные представления о природе и градиенте автоматии. Опыт Станниуса с наложением лигатур.
Занятие № 6.
Занятие № 11.
(20 апреля 2012г.)
Итоговое занятие по теме: Сердечно-сосудистая система.
Занятие № 12.
Физиология больших полушарий. Понятие о высшей нервной деятельности (ВНД).
(8 июня 2012 г.)
1. Эволюционное развитие больших полушарий. Последствия удаления больших полушарий у животных различных видов.
2. Общий план строения коры больших полушарий. Локализация функций в коре больших полушарий. Понятие о полях Бродмана.
3. Методы исследования функций коры больших полушарий.
а) электрофизиологические методики исследования
б) тесты-опросники (клинический опрос)
в) клинико-анатомический
- Понятие о первичных (проекционных), вторичных и ассоциативных зонах. Локализация проекционных зон в различных долях коры больших полушарий у человека, их функциональная роль.
5. Учение И.П. Павлова о высшей нервной деятельности (ВНД):
а) формы приспособительной деятельности животных и человека (безусловные рефлексы, инстинкты и условные рефлексы).
б) значение учения о ВНД для философии, биологии, психологии, педагогики и медицины.
в) понятие об условном рефлексе, его значение и отличие от
безусловного рефлекса.
г) методика образования условных рефлексов, зависимость скорости
образования рефлекса от силы безусловного раздражителя, на базе
которого он вырабатывается.
д) механизм образования условного рефлекса.
6. Классификация условных рефлексов.
7. Торможение в коре больших полушарий, его значение.
8. Виды торможения в коре:
безусловное (внешнее) торможение: постоянный и гаснущий тормоз;
запредельное ( охранительное) торможение;
условное (внутреннее) торможение и его виды (угасательное,
дифференцировочное, условный тормоз, запаздывательное).
9. Движение и взаимоотношения нервных процессов в коре больших
полушарий: иррадиация и концентрация возбуждения, явление индукции.
Занятие № 19.
План
практических занятий по нормальной физиологии для студентов 1
курса стоматологического факультета на весенний семестр 2011 – 2012 учебного года
Занятие № 1.
Введение в физиологию. Основные характеристики живого организма.
(10 февраля 2012 г.)
Занятие № 2.
фазы потенциала действия, механизм возникновения. Восстановительный период. Явление аккомодации возбудимой ткани.
Потенциал действия. Если участок нервного или мышечного волокна подвергнуть действию достаточно сильного раздражителя (например, толчка электрического тока), в этом участке возникает возбуждение, одним из наиболее важных проявлений которого служит быстрое колебание МП, называемое потенциалом действия (ПД)
Причиной возникновения ПД является изменение ионной проницаемости мембраны. В состоянии покоя, как уже говорилось, проницаемость мембраны для К+превышает натриевую проницаемость. Вследствие этого поток положительно заряженных ионов из протоплазмы наружу превышает противоположный поток Na+. Поэтому мембрана в покое снаружи заряжена положительно.
При действии на клетку раздражителя проницаемость мембраны для ионов Na+ резко повышается, и в конечном итоге становится примерно в 20 раз больше проницаемости для К+. Поэтому поток ионов Na+в клетку начинает значительно превышать направленный наружу поток К+. Ток Na+достигает величины +150 мв. Одновременно несколько уменьшается выход К+из клетки. Все это приводит к извращению (реверсии) МП, и наружная поверхность мембраны становится заряженной электро отрицательно по отношению к внутренней поверхности. Указанный сдвиг и регистрируется в виде восходящей ветви пика ПД (фаза деполяризации).
При внутриклеточном отведении можно обнаружить, что поверхность возбужденного участка на очень короткий интервал, измеряемый тысячными долями секунды, становится заряженным электроотрицательно по отношению к соседнему, покоящемуся участку, т.е. при возбуждении происходит т.н. «перезарядка мембраны». Точные измерения показали, что амплитуда ПД на 30-50 мв превышает величину МП. Причина этого состоит в том, что при возбуждении происходит не просто исчезновение ПП, а возникает разность потенциалов обратного знака, в результате чего наружная поверхность мембраны становится заряженной отрицательно по отношению у ее внутренней стороне.
Потенциал действия протекает фазно. Временной ход потенциала действия включает четыре последовательных этапа: локальный ответ, деполяризацию, реполяризацию и следовые потенциалы (рис. 2). В ПД принято различать его пик (т.н. спайк — spike) и следовые потенциалы. Пик ПД имеет восходящую и нисходящую фазы. Перед восходящей фазой регистрируется более или менее выраженный т.н. местный потенциал , или локальный ответ. Поскольку во время восходящей фазы исчезает исходная поляризация мембраны, ее называют фазой деполяризации; соответственно нисходящую фазу, в течение которой поляризация мембраны возвращается к исходному уровню, называется фазой реполяризации. Продолжительность пика ПД в нервных и скелетных мышечных волокнах варьирует в пределах 0,4-5,0 мсек. При этом фаза реполяризации всегда продолжительнее.
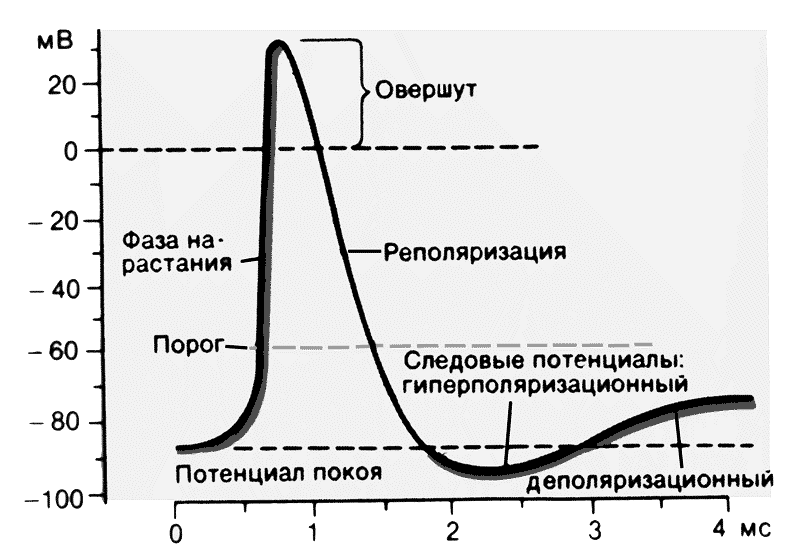
Рис. 2. Фазы и временной ход потенциала действия.
Кроме пика, в ПД различают два следовых потенциала — следовую деполяризацию (следовой отрица-тельный потенциал) и следовую гиперполяризацию (следовой положи-тельный потенциал. Амплитуда этих потенциалов не превышает нескольких милливольт, а длительность варьирует от нескольких десятков до сотен миллисекунд. Следовые потенциалы связаны с восстановительными процессами, развивающимися в мышцах и нерве после окончания возбуждения.
Промежуток времени, в течение которого сохраняется активное состояние в виде ПД, неодинаков в разных возбудимых структурах. В нейронах он составляет около 1 мс, в волокнах скелетных мышц – 10 мс, в миокарде достигает 200–250 мс.
Левое крыло графической записи ПД, отражающее изменение потенциала в электроположительную сторону называется деполяризацией. Область электроположительности носит название овершута, правое крыло ПД, свидетельствующее о восстановлении исходного поляризованного состояния мембраны принято называть реполяризацией. Часто, но не всегда возвращение ПД к исходному уровню в состоянии покоя происходит с наличием фаз в форме так называемых следовых потенциалов. Следовые потенциалы неодинаковы в мышцах и нервах. В волокнах скелетных мышц фаза реполяризации очень замедлена. Примерно через 1 мс после начала ПД наблюдается отчетливый перегиб крыла реполяризации – это следовая деполяризация. В нейроне чаще всего кривая реполяризации быстро пересекает уровень МПП и на некоторое время потенциал мембраны становится более электроотрицательным, чем МП. Это явление называют следовой гиперполяризацией.
Повышение проницаемости мембраны для ионов Na+продолжается в нервных клетках очень короткое время. Связано оно с кратковременным открытием т.н. Na+-каналов (точнее, заслонок М в этих каналах), которое затем сменяется срочным закрытием Na+-пор с помощью т.н. Н-ворот. Этот процесс называется натриевой инактивацией. В результате поток Na в клетку прекращается.
Наличие специальных Na- и К- каналов и сложного механизма запирания и открытия ворот изучено биофизиками достаточно хорошо. Показано, что существуют избирательные механизмы, регулирующие те или иные каналы. Например, яд тетродотоксин блокирует только Na-поры, а тетраэтиламмоний — только К-поры. Показано, что у некоторых клеток возникновение возбуждения связано в изменением проницаемости мембраны для Са++, в других — для Mg+. Исследования механизмов изменения проницаемости мембран продолжаются.
В результате Na-инактивации и одновременного увеличения К- проницаемости происходит усиленный выход положительных ионов К+ из протоплазмы во внешний раствор. В итоге этих двух процессов происходит восстановление поляризованного состояния мембраны (реполяризация) , и наружная ее поверхность вновь приобретает положительный заряд. В дальнейшем происходят процессы восстановления нормального ионного состава клетки и необходимого градиента концентрации ионов за счет активизации деятельности Na-К-насоса. В результате повышения проводимости резко возрастает поток катионов Na+, поэтому отрицательный заряд в клетке вблизи внутренней стороны поверхности мембраны также резко уменьшается вплоть до преобладания положительных зарядов. В результате происходит изменение знака потенциала, достигающего +30 мВ. После этого проводимость мембраны дляNa+также резко снижается.
Для нормального протекания ПД играет существенную роль и изменение проводимости мембраны для K+, которая начинает возрастать позже возрастания проводимости дляNa+. Увеличение относительно медленного выходаK+из клетки в фазу снижения проводимости дляNa+вызывает реполяризацию мембраны.
Таким образом, в живой клетке существуют два различных типа движения ионов через мембрану. Один из них осуществляется по градиенту концентрации ионов и не требует затраты энергии, поэтому его называют пассивным транспортом. Он ответственен за возникновение МП и ПД и ведет в конечном итоге к выравниванию концентраций ионов по обе стороны клеточной мембраны. Второй тип движения ионов через мембрану, осуществляющийся против концентрационного градиента, состоит в «выкачивании» ионов Na+ из протоплазмы и «нагнетании» ионов К+ внутрь клетки. Этот тип ионного транспорта возможет лишь при условии затраты энергии — это активный транспорт. Он является результатом работы специальных ферментных систем (т.н. насосов), и благодаря ему восстанавливается исходная разность концентраций, необходимая для поддержания МП.
Условия возникновения возбуждения. Для возникновения ПД необходимо, чтобы под влиянием какого-либо раздражителя произошло повышение ионной проницаемости мембраны возбудимой клетки. Однако, возбуждение возможно лишь при условии, если действующий на мембрану агент имеет некоторую минимальную (пороговую) величину, способную изменить мембранный потенциал (МП, или Ео ) до некоторого критического уровня (Ек, критический уровень деполяризации). Стимулы, сила которых ниже пороговой величины, называются подпороговыми, выше — надпороговыми. Показано, что пороговая сила, необходимая для возникновения возбуждения при внутриклеточном микроэлектроде равна 10 -7 — 10-9 А.
Таким образом, главным условием для возникновения ПД является следующее: мембранный потенциал должен стать равным или меньше критического уровня деполяризации ( Ео <= Eк)
Инактивация Na+-системы.Na+-системой обозначают механизм, позволяющий в течение нескольких долей миллисекунды многократно (до 20 раз) увеличить проводимость клеточной мембраны для Na+. Достигнув пикового значения, примерно через 0,5 мс проводимость мембраны для Na+ начинает снижаться. Быстрое снижение проводимости для Na+ называют инактивацией Na+-системы. В основе инактивации Na+-системы лежит переход в инактивационное состояние потенциалзависимых Na+-каналов. Поэтому скорость и степень снижения проводимости потенциалзависимы. Это означает, что чем больше отличается потенциал мембраны от мембранного потенциала покоя в сторону электроположительности, тем сильнее инактивирована Na+-система. Поэтому деполяризация мембраны вызывает снижение тока Na+ внутрь клетки. С одной стороны, это свидетельствует о том, что усиление тока Na+ само себе служит причиной его быстрого последующего снижения и начала развития реполяризации. С другой стороны, это означает, что если исходный потенциал клетки выше потенциала покоя на 20–30 мВ, то Na+-система полностью инактивирована и никакая последующая деполяризация уже не может активировать ее, т.е. вызвать резкое увеличение проводимости для Na+ и генерацию ПД.
Кафедра НОРМАЛЬНОЙ ФИЗИОЛОГИИ С БИОФИЗИКОЙ — КиберПедия
ВОПРОСЫ ДЛЯ ПОДГОТОВКИ К ПРАКТИЧЕСКИМ ЗАНЯТИЯМ ДЛЯ СТУДЕНТОВ ЛЕЧЕБНОГО, ПЕДИАТРИЧЕСКОГО И ФАРМАЦЕВТИЧЕСКОГО ФАКУЛЬТЕТОВ
Нормальная физиология является важнейшей фундаментальной медико-биологической дисциплиной. Её изучение является необходимым компонентом подготовки будущего врача и фармацевта, без знания и глубокого понимания этой дисциплины их профессиональная деятельность невозможна.
Перечень вопросов для подготовки к практическим занятиям по нормальной физиологии соответствует требованиям ФГОС по специальностям 060101 – лечебное дело, 060103 – педиатрия и 060301 – фармация.
Пособие состоит из двух частей: в первой представлены контрольные вопросы для студентов лечебного и педиатрического факультета, во второй – для студентов фармацевтического факультета.
В конце списка вопросов для каждой темы указывается список литературных источников для изучения материала. В качестве источников литературы выбраны издания, являющиеся основными в УМК по соответствующей дисциплине. Вопросы и литературные источники для студентов педиатрического факультета выделены жирным курсивом.
Часть I
КОНТРОЛЬНЫЕ ВОПРОСЫ К ПРАКТИЧЕСКИМ ЗАНЯТИЯМ ДЛЯ СТУДЕНТОВ ЛЕЧЕБНОГО И ПЕДИАТРИЧЕСКОГО ФАКУЛЬТЕТОВ
Контрольные вопросы к занятию № 1 на тему:
ВВЕДЕНИЕ В ФИЗИОЛОГИЮ. ОСНОВНЫЕ ХАРАКТЕРИСТИКИ ЖИВОГО ОРГАНИЗМА
1. Предмет и метод физиологии.
2. Связь физиологии с другими дисциплинами, ее значение для медицины.
3. Аналитический и системный подходы в изучении физиологических процессов и функций. Краткая характеристика этапов развития нормальной физиологии: эмпирического, анатомо-физиологического, функционального. Становление и развитие физиологии в XIX – XX вв. Вклад зарубежных и отечественных физиологов в развитие мировой физиологической науки.
4. Понятие о физиологических функциях и процессах клеток, тканей, органов, систем и организма в целом.
5. Основные физиологические функции живого организма (обмен веществ и энергии, рост, размножение, пищеварение, дыхание). Понятие о гомеостазе.
6. Показатели, характеризующие нормальное состояние организма (температура тела, частота и характер пульса, частота дыхания, артериальное давление, осмотическое давление крови, рН и др.).
7. Возрастные периоды развития детского организма.
8. Понятие о регуляции функций. Основные принципы формирования и регуляции физиологических функций: по отклонению, возмущению, прогнозированию. Уровни и механизмы (нервный, гуморальный) регуляции функций. Представление о саморегуляции постоянства внутренней среды организма.
9. Общие свойства возбудимых тканей: раздражимость и возбудимость. Виды раздражителей.
10. Порог раздражения (реобаза). Полезное время действия тока. Хронаксия. Кривая зависимости силы и времени действия тока.
11. Изменение возбудимости при возбуждении: рефрактерная фаза (абсолютная и относительная), фаза повышенной (супернормальной) и фаза пониженной (субнормальной) возбудимости.
Литература:
1.Учебник «Медицинская физиология» // А. Гайтон, Д. Холл, – М., 2008, С. 3 – 10.
2.Учебник «Физиология человека» под ред. В.М. Покровского, Г.Ф. Коротько, – М.: Медицина, 2003, С. 17 – 39.
3.Лекционные материалы
4.Методическое пособие по возрастной физиологии человека для студентов педиатрического факультета. Ярославль, 1977, С. 3 – 6.
Контрольные вопросы к занятию № 2 на тему:
ФИЗИОЛОГИЯ КЛЕТОЧНЫХ МЕМБРАН. ЭЛЕКТРИЧЕСКИЕ ЯВЛЕНИЯ В НЕРВНО-МЫШЕЧНОЙ СИСТЕМЕ
1. Клеточные мембраны, их виды (цитоплазматическая, аппарата Гольджи, митохондрий, саркоплазматического ретикулума, лизосом, ядерная оболочка и др.), их роль. Химический состав цитоплазматической мембраны. Мембранные белки (интегральные, периферические, транспортные), их роль.
2. Свойства мембран. Функции мембран.
3. Рецепторы, их классификация: по локализации (мембранные, ядерные), механизму развития процессов (ионо- и метаботропные), по скорости приема сигнала (быстрые, медленные, по роду воспринимающих веществ.
4. Ионотропные рецепторы, метаботропные рецепторы и их разновидности. Системы вторичных посредников действия метаботропных рецепторов (цАМФ, цГМФ, инозитол-3- фосфат, диацилглицерол, ионы Са++).
5. Ионные каналы, их строение. Классификация ионных каналов.
6. Токи покоя (мембранный потенциал, ток повреждения, метаболический ток).
7. Происхождение мембранного потенциала. Значение ионной асимметрии во внеклеточной и внутриклеточной жидкости. Ионные градиенты клетки, их механизмы. Уравнение Нернста.
8. Ионные насосы (АТФ-азы): K+-Na+-евая, Ca2+-евая (плазмолеммы и саркоплазматического ретикулума), H+-K+-обменник.
9. Современные представления о процессе возбуждения. Локальный ответ. Критический уровень деполяризации при действии раздражителей. Понятие о поляризации мембраны, ее деполяризации, реверсии, реполяризации и гиперполяризации.
10. Потенциал действия и его фазы. Условия регистрации двухфазного и однофазного тока действия.
11. Происхождение компонентов кривой однофазного тока действия, их продолжительность. Мембранно-ионная теория происхождения тока действия.
12. Соотношение фаз возбудимости с фазами потенциала действия.
Литература
1. Учебник «Медицинская физиология»// А. Гайтон, Д. Холл, – М., 2008, С. 11 — 18, 47 — 76.
2. Учебник «Физиология человека» под ред. В.М. Покровского, Г.Ф.
Коротько, – М.: Медицина, 2003, С. 39 – 58.
3. Лекционные материалы.
Контрольные вопросы к занятию № 3 на тему:
ФИЗИОЛОГИЯ НЕРВОВ И НЕРВНО-МЫШЕЧНОГО СИНАПСА
1. Значение нервной системы в механизмах регуляции функций в организме. Функция нервов.
2. Строение периферических нервных волокон. Миелинизация нервных волокон в онтогенезе.
3. Физиологические свойства нервных волокон (возбудимость и проводимость).
4. Механизм проведения возбуждения в мякотных и безмякотных нервных волокнах.
5. Классификация нервных волокон по их диаметру, скорости проведения возбуждения и особенностям потенциала действия (группы А, В и С). Возрастные изменения скорости проведения возбуждения.
6. Законы проведения возбуждения в нервах (закон анатомической и физиологической целостности нервного волокна, закон двухстороннего проведения возбуждения и закон изолированного проведения возбуждения).
7. Нервно-мышечный синапс, структура и свойства, медиатор. Понятие о холинорецепторе.
8. Механизм передачи возбуждения с нерва на мышцу, Потенциал концевой пластинки (ПКП) и его свойства. Значение холинэстеразы.
9. Возникновение потенциала действия в мышечном волокне.
10.Понятие о лабильности.
Литература:
1. Учебник «Медицинская физиология» // А.Гайтон, Дж. Холл, – М. 2008, С. 60 – 75, 90 – 97.
2. Учебник «Физиология человека» под ред. В.М. Покровского, Г.Ф. Коротько, – М.: Медицина, 2003, С. 58 – 74.
3. Лекционные материалы.
4. Методическое пособие по возрастной физиологии человека для студентов педиатрического факультета. Ярославль, 1977, С. 58 –59
Контрольные вопросы к занятию № 4 на тему:
ФИЗИОЛОГИЯ МЫШЦ. МЕХАНИЗМ МЫШЕЧНОГО СОКРАЩЕНИЯ
1. Физиологическая роль мышечной системы. Классификация мышц.
2. Морфо-функциональные особенности исчерченных мышц.
3. Ультраструктура миофибрилл: тонкие и толстые протофибриллы, их взаимное расположение в саркомере.
4. Сократительные белки (миозин и актин) и регуляторные белки (тропомиозин и тропонин) в составе протофибрилл, их свойства.
5. Понятие об L-системе продольных трубочек саркоплазматического ретикулума, их значение как депо Са2+. Т-система поперечных трубочек мембраны миофибрилл и ее взаимодействие с L-системой.
6. Теория мышечного сокращения и расслабления (Хаксли, Катц).
7. Биоэлектрические, химические и тепловые процессы в мышцах.
8. Электромеханическая связь возбуждения и сокращения мышцы.
Итоговое занятие по теме: Кровь. Дыхание
План
практических занятий по нормальной физиологии для студентов 2
курса лечебного факультета на осенний семестр 2011 – 2012 учебного года
Занятие № 1.
Введение в физиологию. Основные характеристики живого организма.
(5 — 10 сентября 2011 г.)
Занятие № 2.
Физиология клеточных мембран. Электрические явления в нервно-мышечной системе
(12 — 17 сентября 2010 г.)
Занятие № 3.
Физиология нервов и нервно-мышечного синапса
(19-24 сентября 2011 г.)
Занятие № 4.
Физиология мышц. Механизм мышечного сокращения
(26 сентября — 1 октября 2011 г.)
Занятие № 5.
Общая физиология центральной нервной системы. Основные свойства нервных центров
(3 — 8 октября 2011 г.)
Занятие № 6.
Торможение в центральной нервной системе. Физиология спинного мозга
(10 — 15 октября 2011 г.)
Занятие № 7.
Физиология продолговатого, среднего и промежуточного мозга
(17-22 октября 2011 г.)
Занятие № 8.
Физиология ретикулярной формации. Мозжечок и базальные ганглии. Экстрапирамидная и пирамидная системы.
(24 — 29 октября 2011 г.)
Занятие № 9.
Физиология автономной (вегетативной) нервной системы
(31 октября-5 ноября 2011 года)
Занятие № 10.
Итоговое занятие по теме: Центральная нервная система.
(7-12 ноября 2011 года)
Занятие № 11.
Физиология желез внутренней секреции
(14-19 ноября 2011 года)
Занятие № 12.
Форменные элементы крови
(21-26 ноября 2011 г.)
Занятие № 13.
Скорость оседания эритроцитов. Дыхательная функция крови
(28 ноября — 3 декабря 2011 г.)
Занятие № 14.
Гемостаз и свертывание крови. Видовые и индивидуальные свойства крови
(5-10 декабря 2011 г.)
Занятие № 15.
Механизм вентиляции и газообмен в легких
(12 -17 декабря 2011 г.)
Занятие № 16.
Транспорт газов кровью. Регуляция дыхания
(19-24 декабря 2011 г)
Занятие № 17.
Итоговое занятие по теме: Кровь. Дыхание
(26-31 декабря 2011 года)
Контрольные вопросы к занятию № 1 на тему:
Введение в физиологию. Основные характеристики живого организма.
(5 — 10 сентября 2011 г.)
1. Предмет и метод физиологии.
2. Связь физиологии с другими дисциплинами, ее значение для медицины.
3. Аналитический и системный подходы в изучении физиологических процессов и функций. Краткая характеристика этапов развития нормальной физиологии: эмпирического, анатомо-физиологического, функционального. Становление и развитие физиологии в XIX-XX вв. Вклад зарубежных и отечественных физиологов в развитие мировой физиологической науки.
4. Понятие о физиологических функциях и процессах клеток, тканей, органов, систем и организма в целом.
5. Основные физиологические функции живого организма (обмен веществ и энергии, рост, размножение, пищеварение, дыхание). Понятие о гомеостазе.
6. Показатели, характеризующие нормальное состояние организма (температура тела, частота и характер пульса, частота дыхания, артериальное давление, осмотическое давление крови, Рн и др.)
7. Понятие о регуляции функций. Основные принципы формирования и регуляции физиологических функций: по отклонению, возмущению, прогнозированию. Уровни и механизмы (нервный, гуморальный) регуляции функций. Представление о саморегуляции постоянства внутренней среды организма.
8. Общие свойства возбудимых тканей: раздражимость и возбудимость.
9. Виды раздражителей.
10. Порог раздражения (реобаза). Полезное время действия тока. Хронаксия. Кривая зависимости силы и времени действия тока.
11. Изменение возбудимости при возбуждении: рефрактерная фаза (абсолютная и относительная), фаза повышенной (супернормальной) и фаза пониженной (субнормальной) возбудимости.
Контрольные вопросы к занятию № 2 на тему:
Физиология клеточных мембран. Электрические явления в нервно-мышечной системе
(12 — 17 сентября 2010 г.)
1. Клеточные мембраны, их виды (цитоплазматическая, аппарата Гольджи, митохондрий, саркоплазматического ретикулума, лизосом, ядерная оболочка и др.), их роль. Химический состав цитоплазматической мембраны. Мембранные белки (интегральные, периферические, транспортные), их роль.
2. Свойства мембран. Функции мембран.
3. Рецепторы, их классификация: по локализации (мембранные, ядерные), механизму развития процессов (ионо- и метаботропные), по скорости приема сигнала (быстрые, медленные, по роду воспринимающих веществ.
4. Ионотропные рецепторы, метаботропные рецепторы и их разновидности. Системы вторичных посредников действия метаботропных рецепторов ( цАМФ, ц ГМФ, инозитол-3- фосфат, диацилглицерол, ионы Са++).
5. Ионные каналы, их строение. Классификация ионных каналов.
6. Токи покоя (мембранный потенциал, ток повреждения, метаболический ток).
7. Происхождение мембранного потенциала. Значение ионной асимметрии во вне и внутриклеточной жидкости. Ионные градиенты клетки, их механизмы. Уравнение Нернста.
8. Ионные насосы (АТФ-азы): K+-Na+-евая, Ca2+-евая (плазмолеммы и саркоплазматического ретикулума), H+–K+-обменник.
9. Современные представления о процессе возбуждения. Локальный ответ. Критический уровень деполяризации при действии раздражителей. Понятие о поляризации мембраны, ее деполяризации, реверсии, реполяризации и гиперполяризации.
10. Потенциал действия и его фазы. Условия регистрации двухфазного и однофазного тока действия.
11. Происхождение компонентов кривой однофазного тока действия, их продолжительность. Мембранно-ионная теория происхождения тока действия.
12. Соотношение фаз возбудимости с фазами потенциала действия.
Контрольные вопросы к занятию № 3 на тему:
Физиология нервов и нервно-мышечного синапса
(19-24 сентября 2011 г.)
1. Значение нервной системы в механизмах регуляции функций в организме. Функция нервов.
2. Строение периферических нервных волокон. Миелинизация нервных волокон в онтогенезе.
3. Физиологические свойства нервных волокон (возбудимость и проводимость).
4. Механизм проведения возбуждения в мякотных и безмякотных нервных волокнах.
5. Классификация нервных волокон по их диаметру, скорости проведения возбуждения и особенностям потенциала действия (группы А, В и С). Возрастные изменения скорости проведения возбуждения.
6. Законы проведения возбуждения в нервах (закон анатомической и физиологической целостности нервного волокна, закон двухстороннего проведения возбуждения и закон изолированного проведения возбуждения ).
7. Нервно-мышечный синапс, структура и свойства, медиатор. Понятие о холинорецепторе.
8. Механизм передачи возбуждения с нерва на мышцу, Потенциал концевой пластинки (ПКП) и его свойства. Значение холинэстеразы.
9. Возникновение потенциала действия в мышечном волокне.
10. Понятие о лабильности.
Контрольные вопросы к занятию № 4 на тему:
Физиология мышц. Механизм мышечного сокращения
(26 сентября — 1 октября 2011 г.)
1. Физиологическая роль мышечной системы. Классификация мышц.
2. Морфо-функциональные особенности исчерченных мышц.
3. Ультраструктура миофибрилл: тонкие и толстые протофибриллы, их взаимное расположение в саркомере.
4. Сократительные белки (миозин и актин) и регуляторные белки (тропомиозин и тропонин) в составе протофибрилл, их свойства.
5. Понятие об L — системе продольных трубочек саркоплазматического ретикулума, их значение как депо Са++. Т — система поперечных трубочек мембраны миофибрилл и ее взаимодействие с L — системой.
6. Теория мышечного сокращения и расслабления (Хаксли, Катц).
7. Биоэлектрические, химические и тепловые процессы в мышцах.
8. Электромеханическая связь возбуждения и сокращения мышцы.
9. Физические и физиологические свойства мышц.
10. Режимы и виды сокращений мышц (одиночные и тетанические).
11. Одиночное сокращение и его фазы. Изменение возбудимости при возбуждении мышцы.
12. Явление суммации сокращений.
13. Тетанические сокращения: зубчатый и гладкий тетанус.
14. Понятие о лабильности (Н.Е. Введенский). Оптимум и пессимум силы и частоты для сокращения мышцы.
15. Основные отличия в строении и функции скелетных и гладких мышц внутренних органов.
Контрольные вопросы к занятию № 5 на тему:
Читайте также:
Рекомендуемые страницы:
Поиск по сайту
31.Принцип действия однофазного мостового инвертора напряжения.

Рисунок 36.1 Схема однофазного полумостового инвертора
со средней точкой источника питания
Для работы данной схемы необходимо иметь источник питания со средней точкой, которая образуется за счет включения на входе инвертора двух емкостей CФ1иCФ2. Транзисторы в данной схеме работают попарно в противофазе с одинаковой длительностью закрытого и открытого состояния.
При активной нагрузке ток и напряжение
имеют одну и ту же форму (прямоугольных
двухполярных импульсов). К нагрузке на
каждом полупериоде прикладывается
напряжение 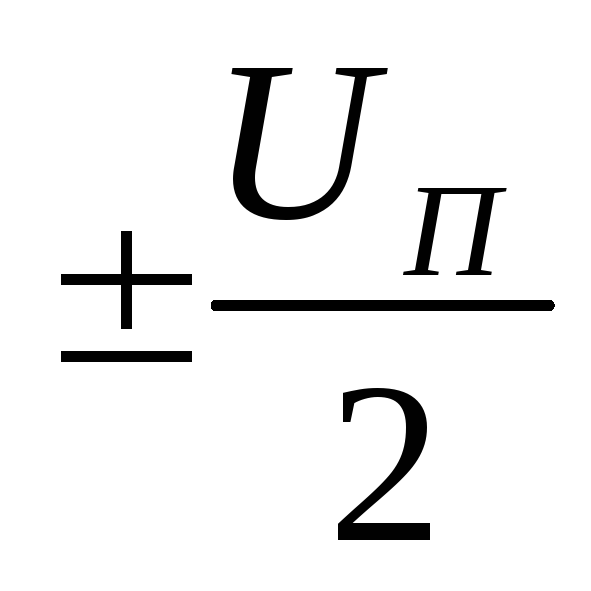 .
.

Рисунок 36.2 Временные диаграммы работы схемы на R—L нагрузку
При наличии индуктивности форма тока
становится апериодической. В момент
времени θ1 открывается
транзисторVT1и
ток нагрузки протекает от источника
питания по цепи+UП – VT1 – RL – ноль источника питания. При этом к
нагрузке прикладывается напряжение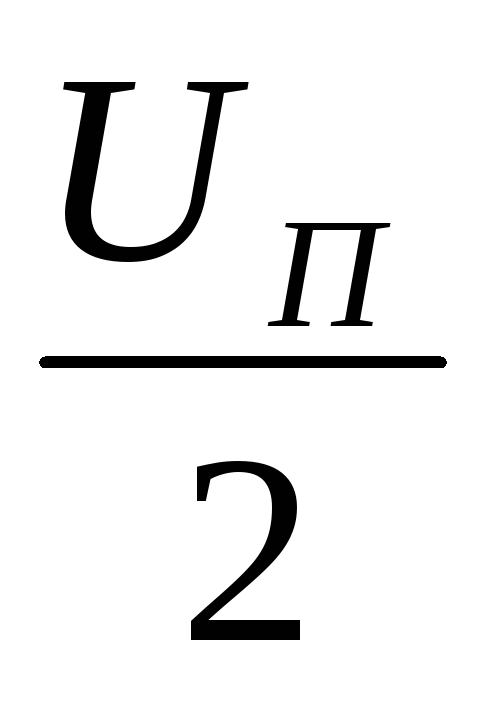 ,
а к транзисторуVT2в прямом и диодуVD2в обратном направлении напряжениеUП.
В момент времениθ2транзисторVT1закрывается и должен открыться транзисторVT2, но из-за действия
э.д.с самоиндукции изменившей свой знак
и стремящейся поддержать ток в цепи
нагрузки откроется диодVD2и ток продолжает протекать в том же
направленииVD2–RL– ноль источника
питания –+CФ2–—UП–VD2. При этом ток в
цепи нагрузки уменьшается, а ток в цепи
источника питания
,
а к транзисторуVT2в прямом и диодуVD2в обратном направлении напряжениеUП.
В момент времениθ2транзисторVT1закрывается и должен открыться транзисторVT2, но из-за действия
э.д.с самоиндукции изменившей свой знак
и стремящейся поддержать ток в цепи
нагрузки откроется диодVD2и ток продолжает протекать в том же
направленииVD2–RL– ноль источника
питания –+CФ2–—UП–VD2. При этом ток в
цепи нагрузки уменьшается, а ток в цепи
источника питания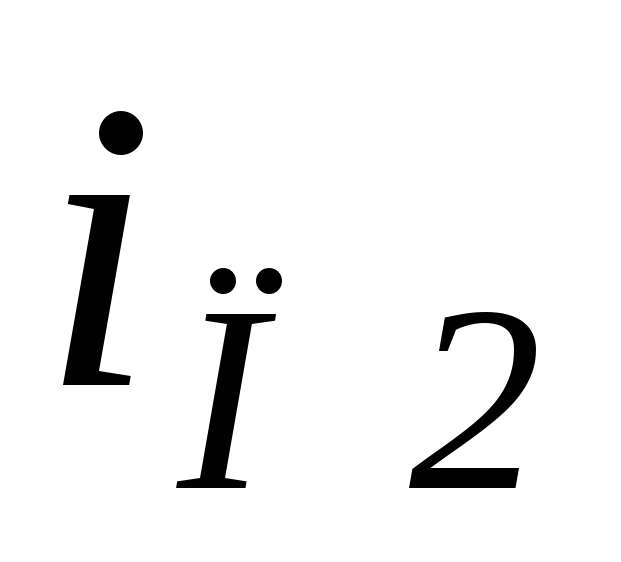 поменял свой знак, то есть на интервалеθ2<θ<θ3ток по нагрузке протекает под действием
ЭДС самоиндукции и происходит возврат
энергии вCФ2.
поменял свой знак, то есть на интервалеθ2<θ<θ3ток по нагрузке протекает под действием
ЭДС самоиндукции и происходит возврат
энергии вCФ2.
В момент времени θ3ток в нагрузке становится равным нулю, диодVD2закрывается, открывается транзисторVT2и ток начинает протекать по цепиVT2 – —UП – CФ2– ноль источника питания –RL–VT2.
Действующее значение ЭДС в нагрузке
равно 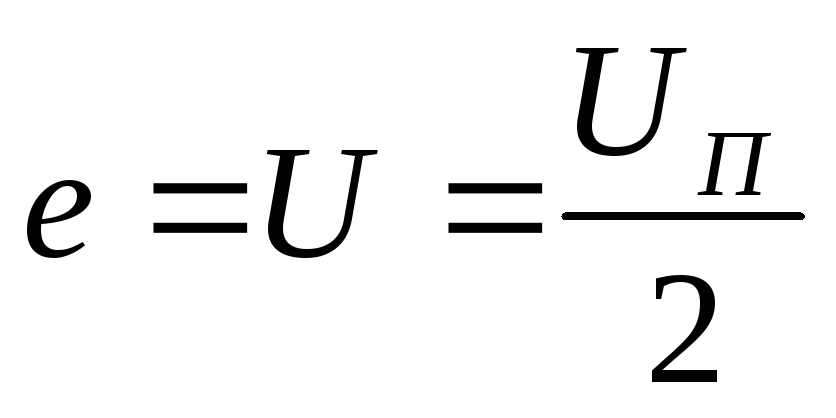 .
.
Недостатки схемы: необходимость источника питания со средней точкой (двух емкостей) и к ключам прикладывается двойное напряжение.
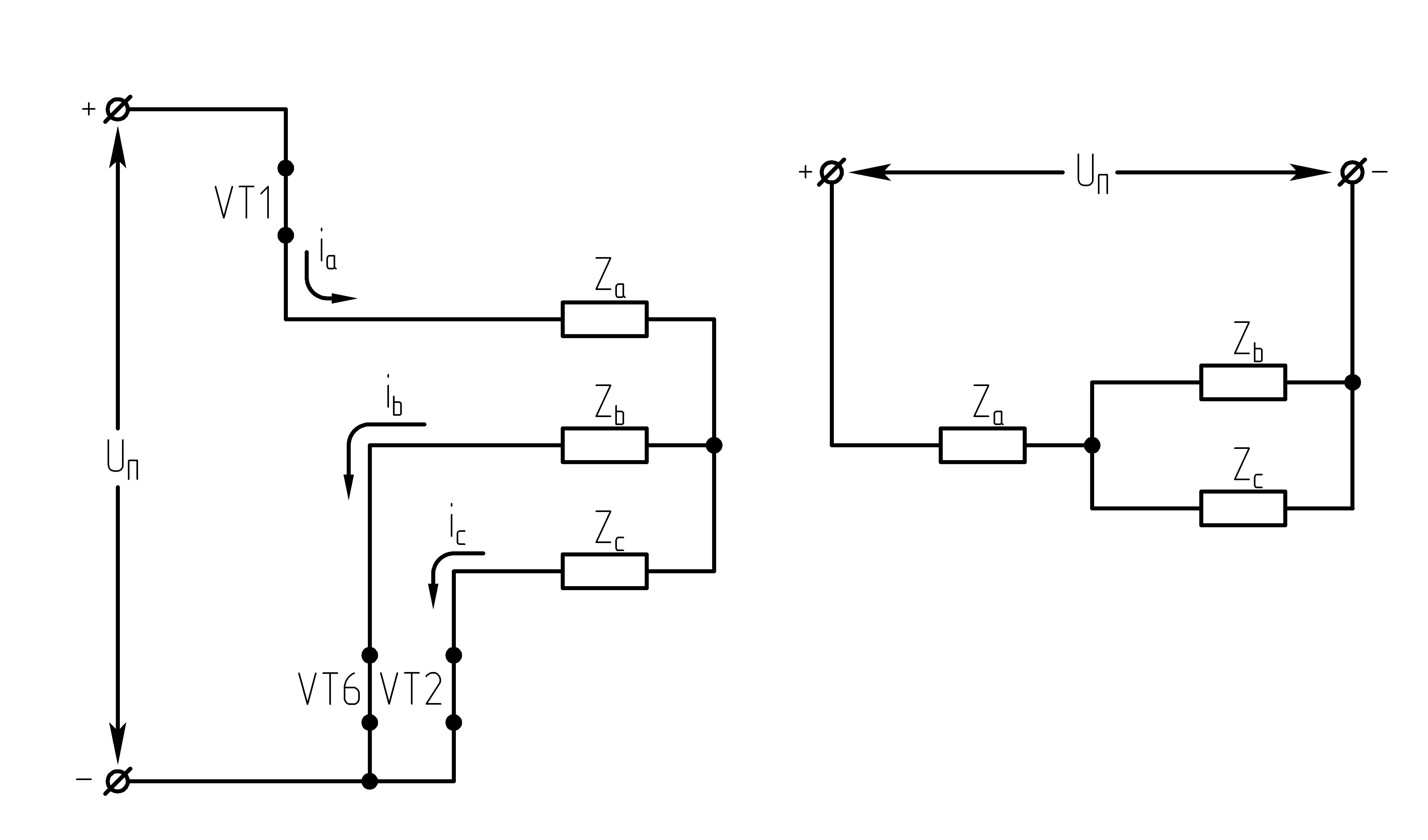
32.Принцип действия трехфазного автономного инвертора напряжения с углом проводимости =180°. Нагрузка активная, соединение-звезда.
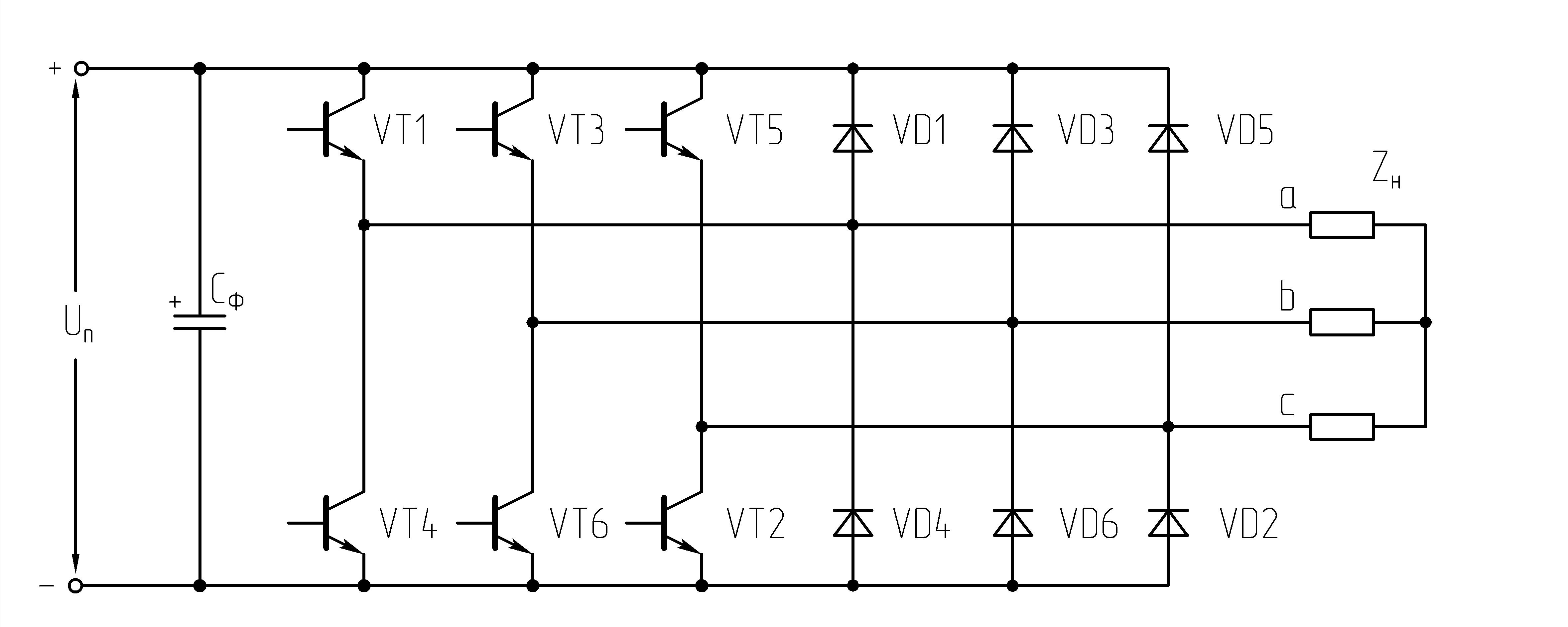
Рисунок 39.1 Схема трехфазного мостового АИН
При λ=1800алгоритм переключения ключей АИН имеет следующий вид:
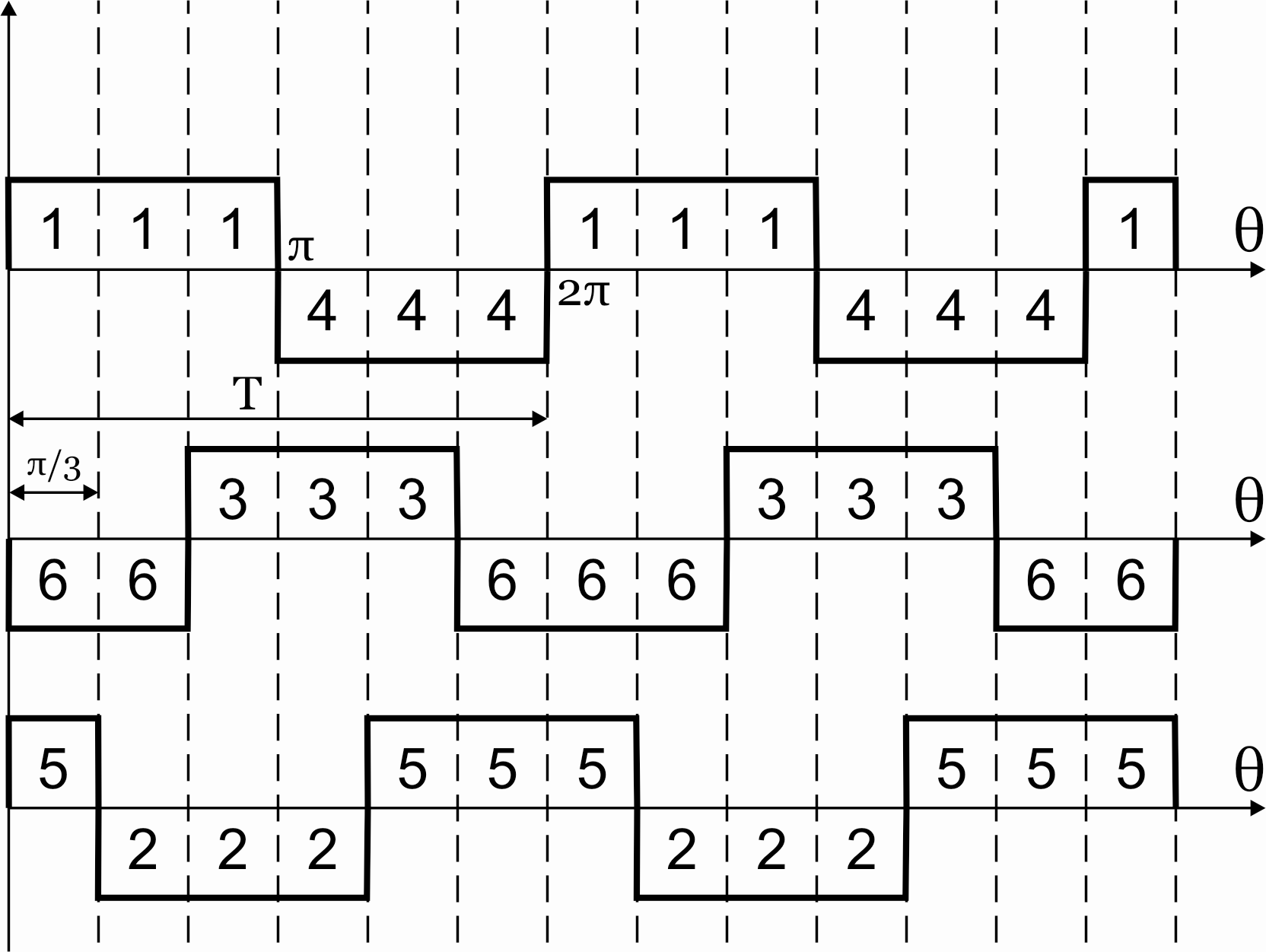
Рисунок 39.2 Алгоритм переключения ключей инвертора при λ=1800
Выполнение данного алгоритма позволяет получить на нагрузке кривую 3-х фазного симметричного несинусоидального переменного напряжения.
Из алгоритма переключения ключей
инвертора следует, что в 3-хфазном
мостовом АИН при λ=1800за
периодТвыходного напряжения
инвертора возможны шесть независимых
сочетаний открытых и закрытых состояний
транзисторов. При этом, в каждый момент
времени открывающие импульсы подаются
одновременно на три транзистора (ключа).
Ток, втекающий черезVT1,VT3иVT5– положительный. 1.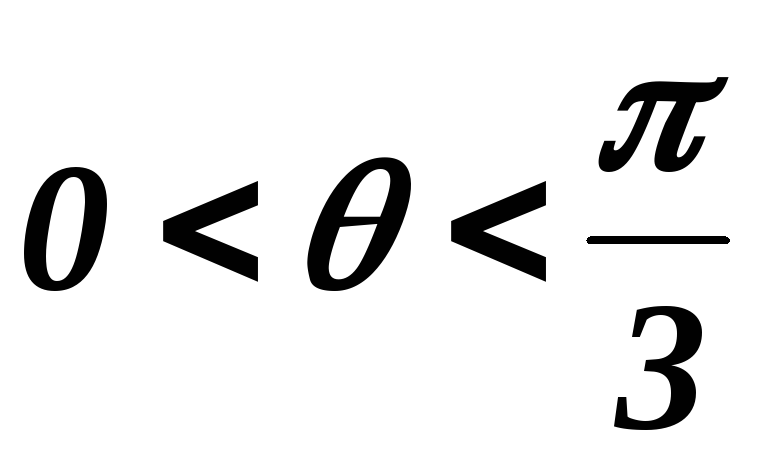
На данном интервале открыты VT1,VT6,VT5. Схема замещения АИН следующая:
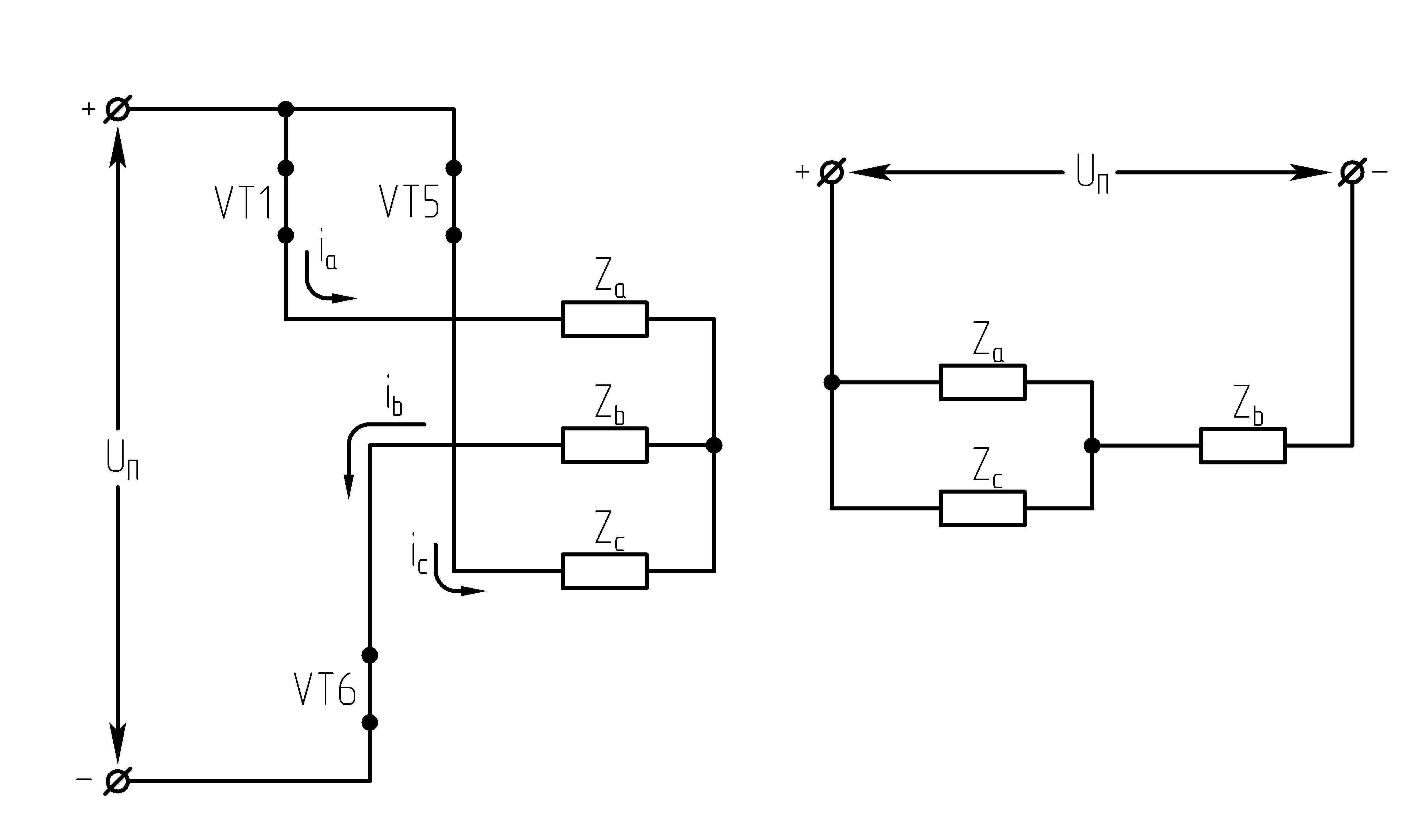
Рисунок
39.3 Схема замещения при 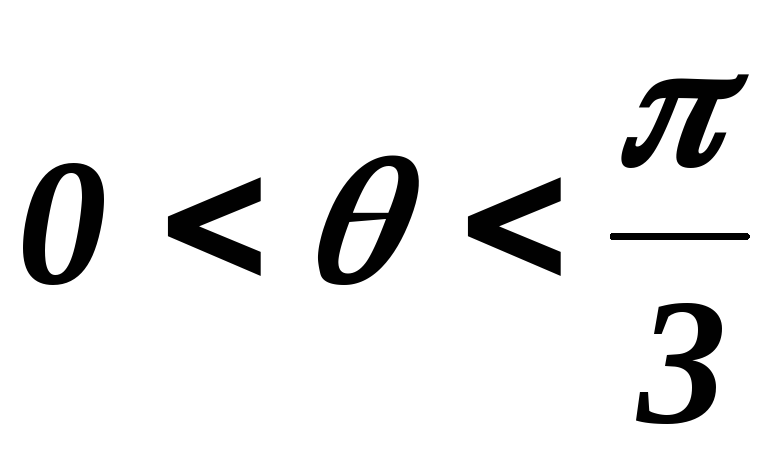
Таким образом, считая, что 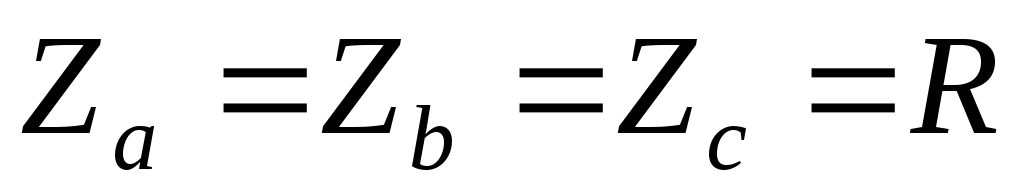
 ;
2.
;
2.
На данном интервале открыты VT1,VT6,VT2. Схема замещения АИН следующая:
Рисунок
39.4 Схема замещения при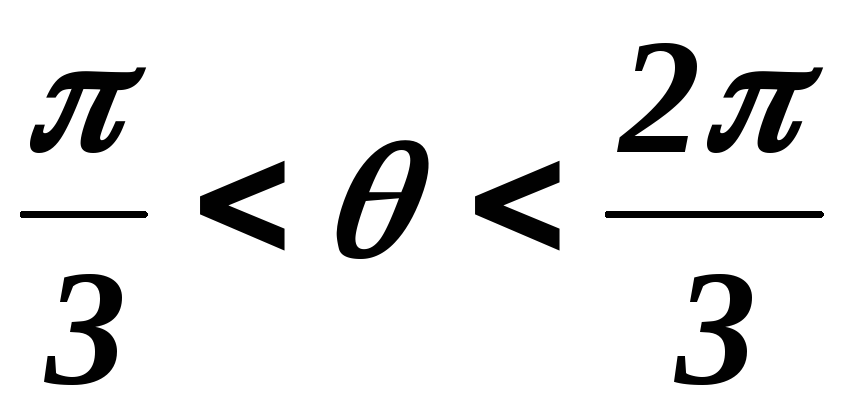
 ;
3.
;
3.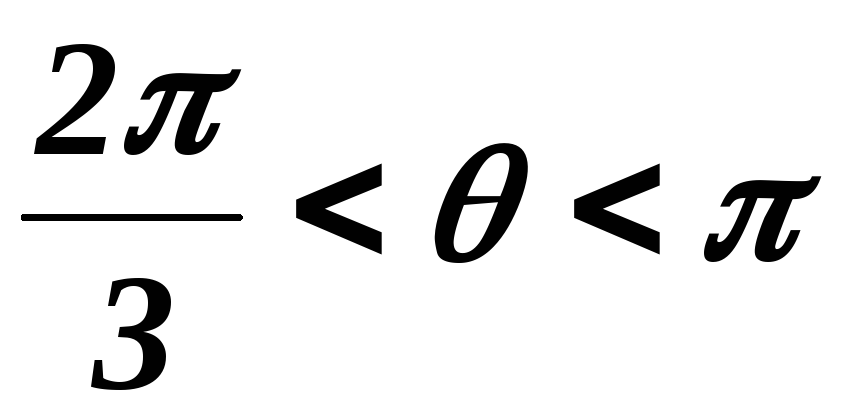
На данном интервале открыты VT1,VT2,VT3. Схема замещения АИН следующая:
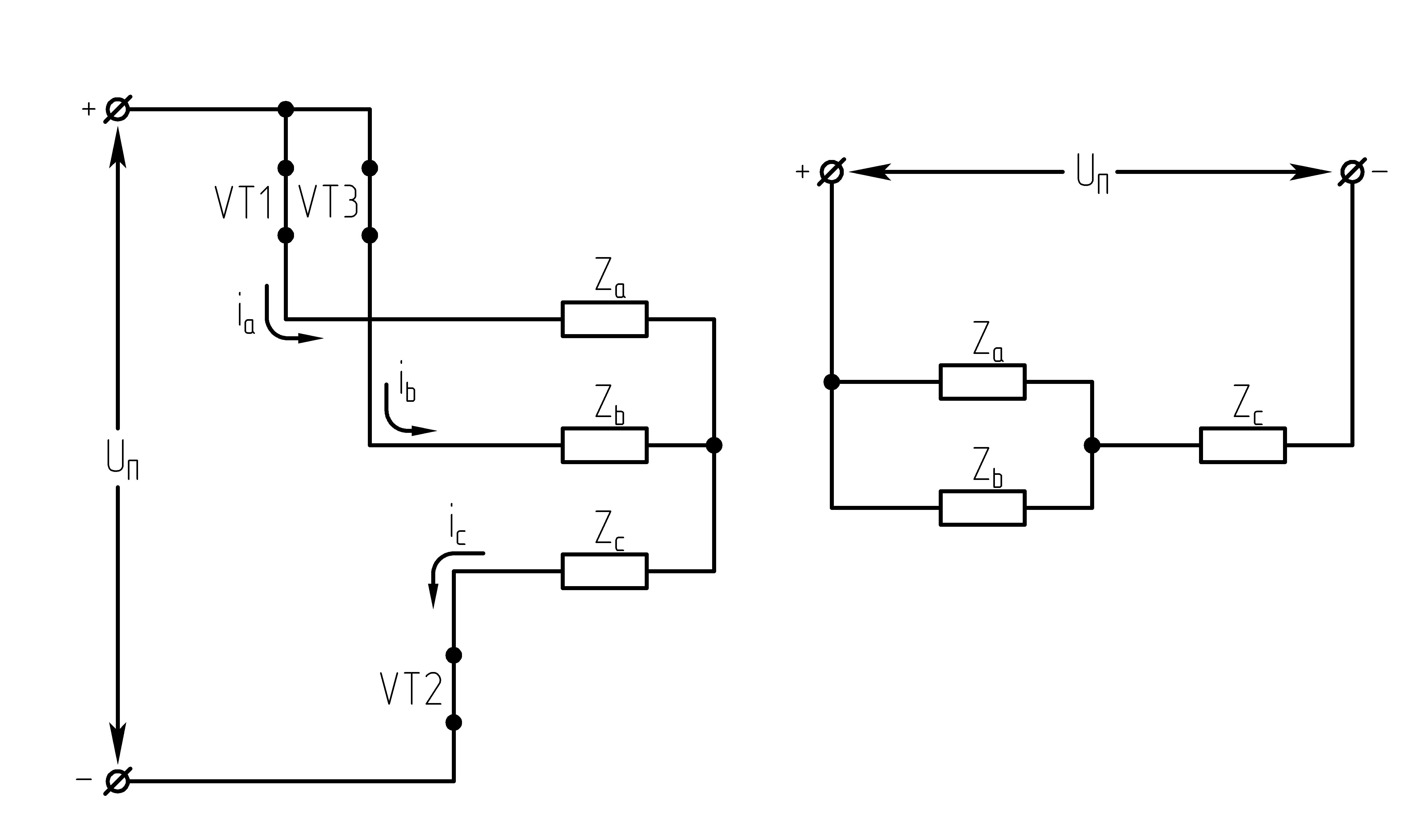
Рисунок
39.5 Схема замещения при 
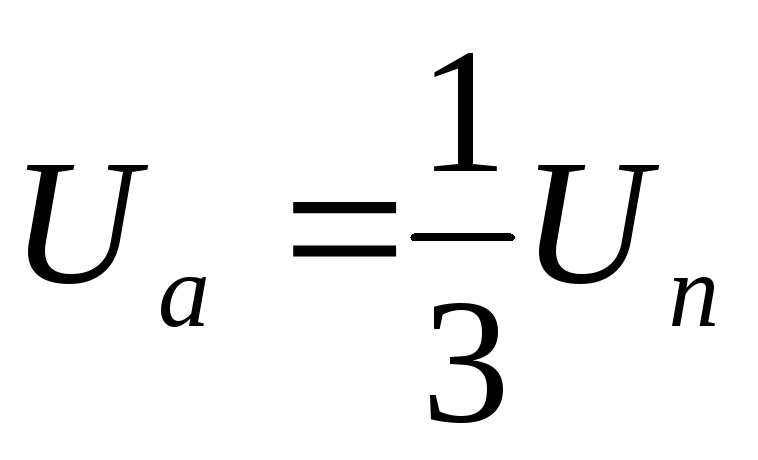 ;
4.
;
4.
На данном интервале открыты VT4,VT3,VT2. Схема замещения АИН следующая:
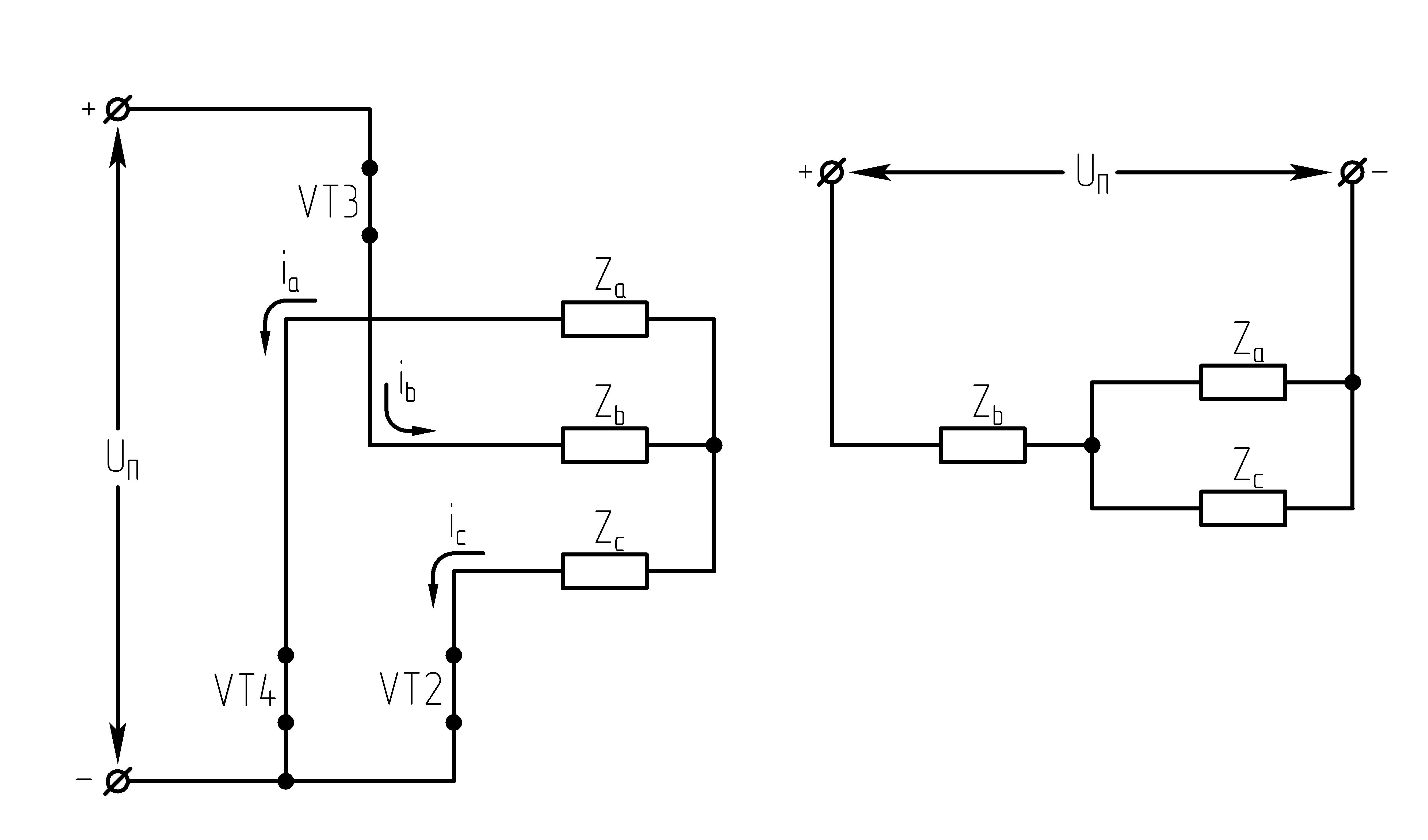
Рисунок 39.6 Схема
замещения при 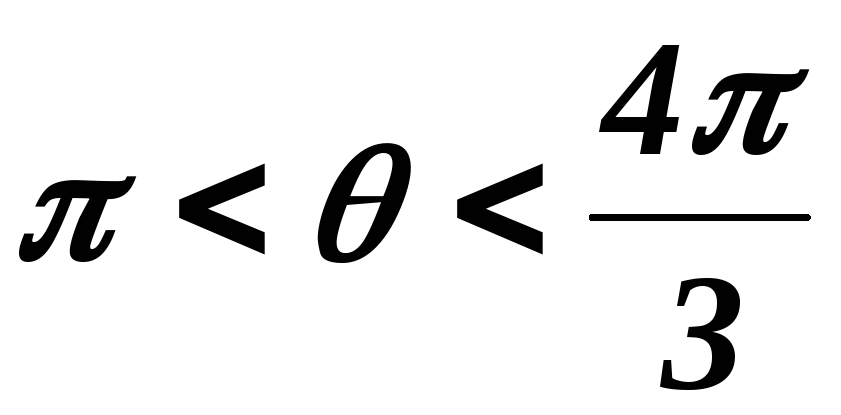
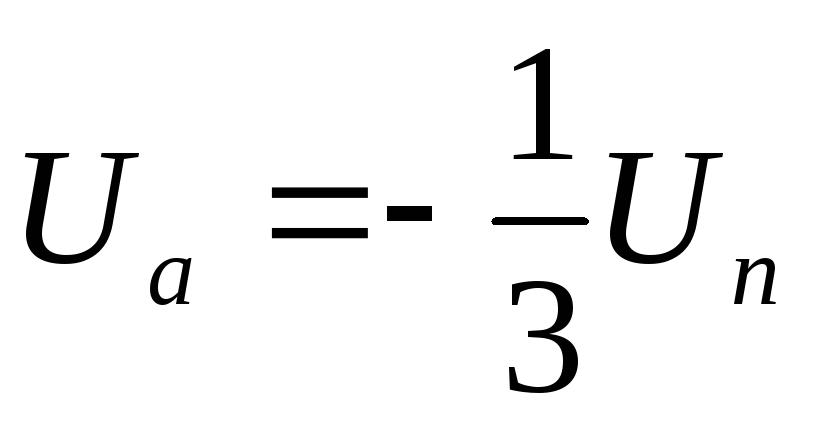 5.
5. 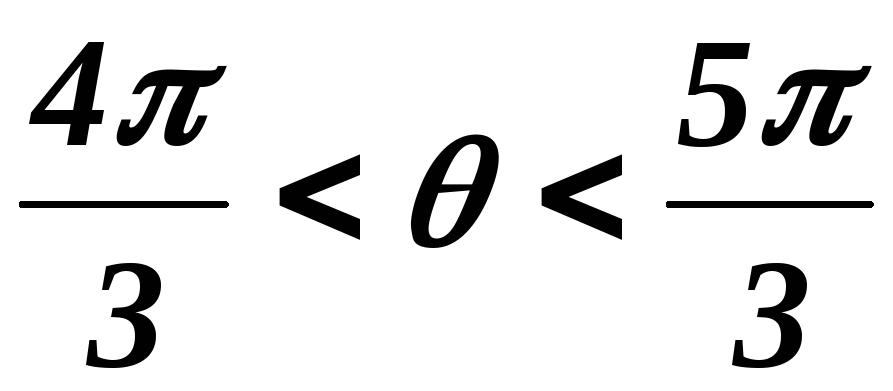
На данном интервале открыты VT5,VT3,VT4. Схема замещения АИН следующая:

Рисунок
39.7 Схема замещения при 
 ;
6.
;
6.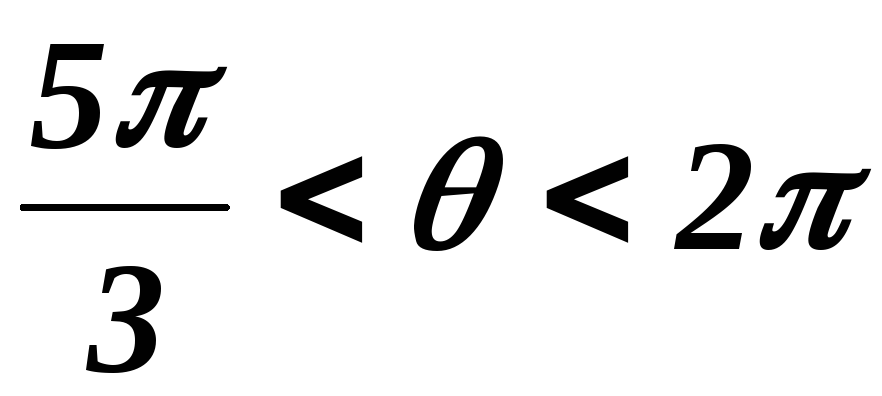
На данном интервале открыты VT4,VT6,VT5. Схема замещения следующая:

Рисунок
39.8 Схема замещения при 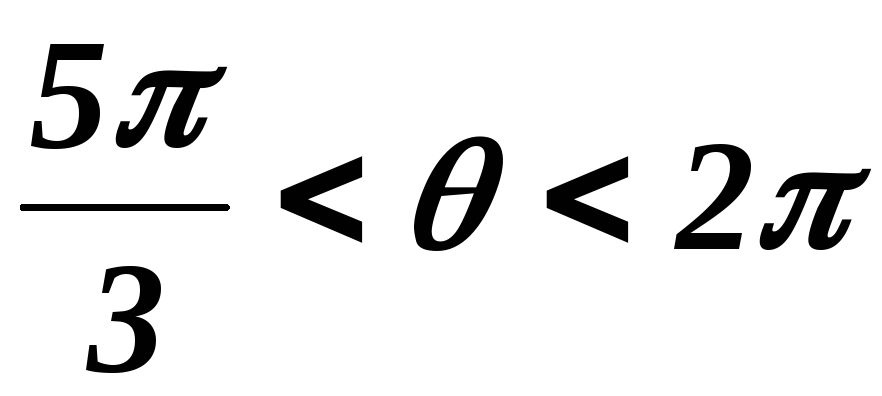
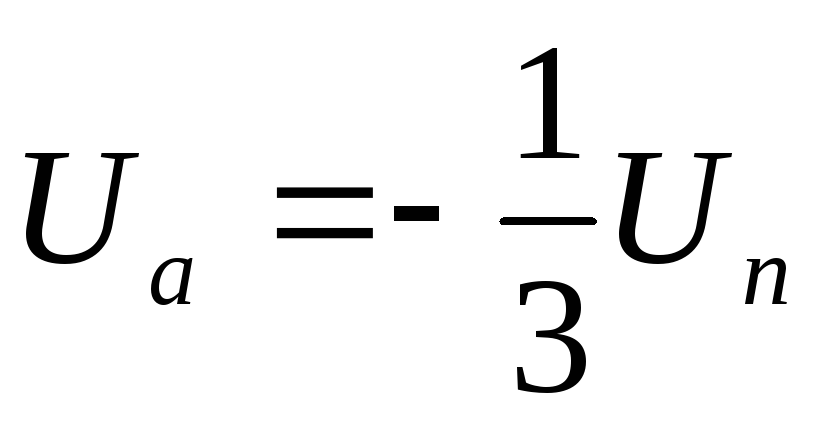
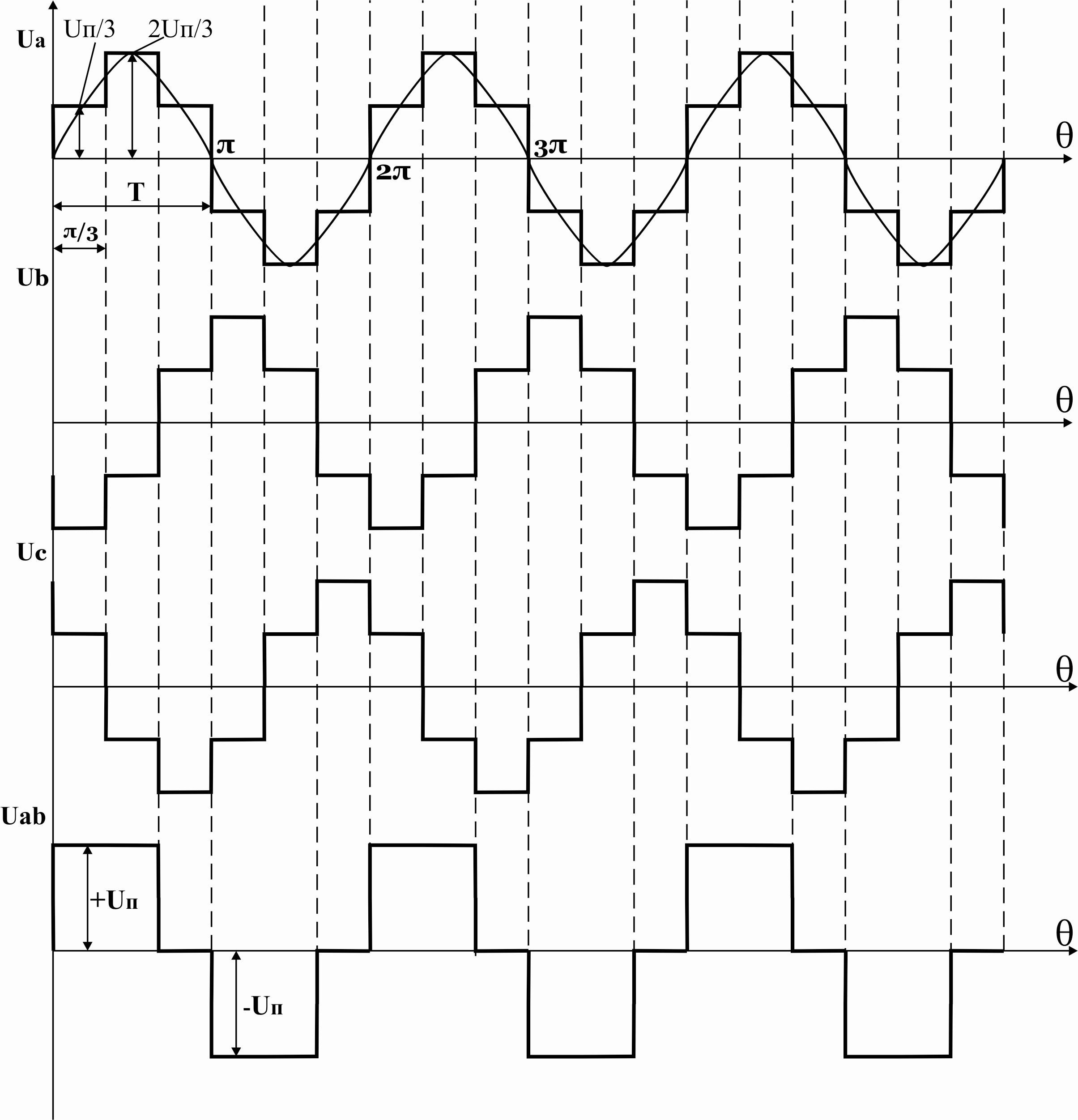
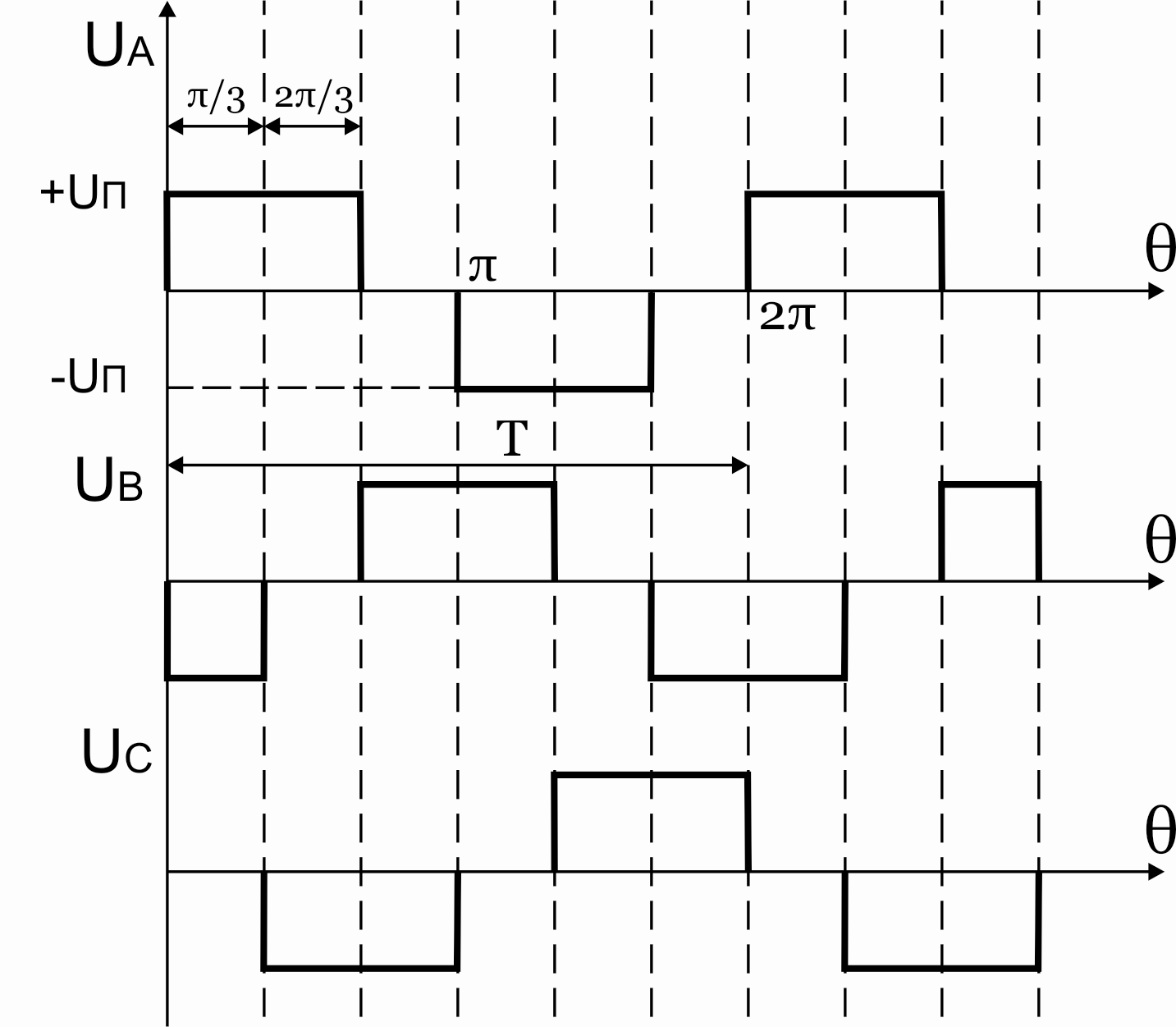
Р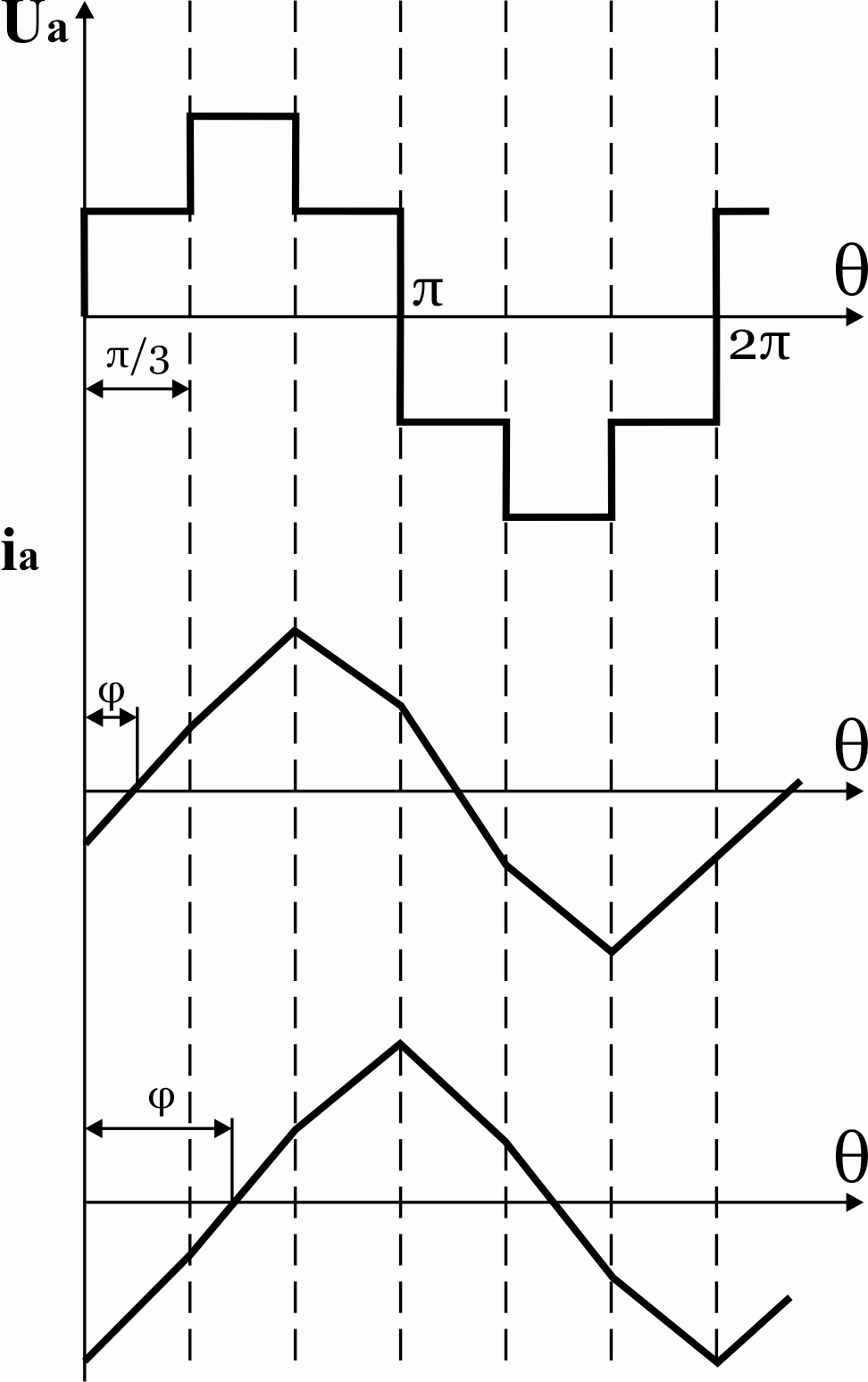 исунок
39.9 Временные диаграммы работы 3-х фазного
исунок
39.9 Временные диаграммы работы 3-х фазного
мостового АИН при λ=1800
Из схем замещения видно , что за период выходного напряжения каждая фаза нагрузки включается либо параллельно другой фазе и последовательно с третьей , либо последовательно с двумя другими фазами, включенными параллельно. Поэтому, к каждой фазе прикладывается напряжение либо Uп/3(-Uп/3), либо 2/3Uп(-2/3Uп).
Фазное напряжение на нагрузке двухполярное шестиступенчатое. При активной нагрузке кривая тока повторяет кривую напряжения.
Трехфазный мостовой АИН λ=1800, соединение нагрузки в треугольник (Δ)
При соединении активной нагрузки в Δ возможны также шесть сочетаний открытых и закрытых состояний транзисторов.

Рисунок 39.10 Схемы замещений
1. 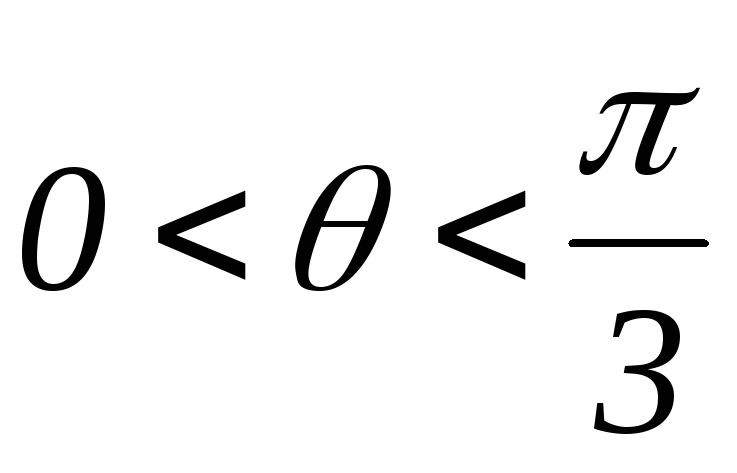 :
открытыVT1,VT6, VT5;
:
открытыVT1,VT6, VT5;
2. 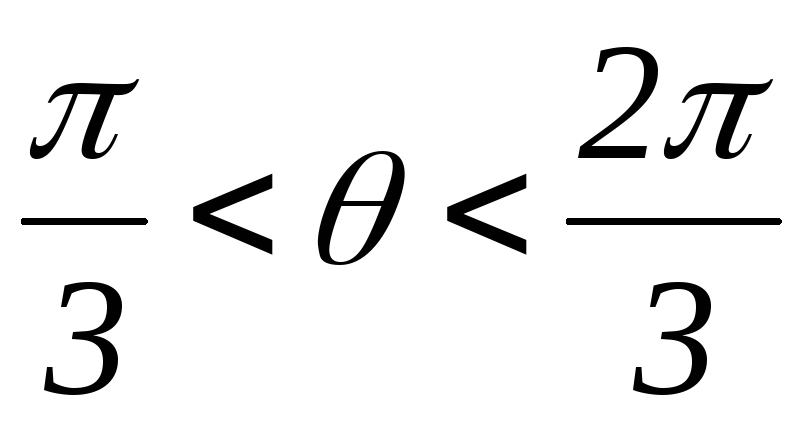 :
открытыVT1,VT2, VT6;
:
открытыVT1,VT2, VT6;
3. 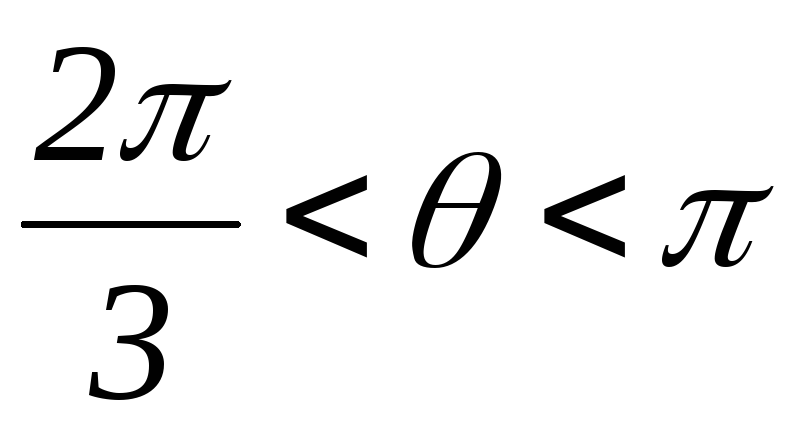 :
открытыVT1,VT3, VT2.
:
открытыVT1,VT3, VT2.
При соединении активной нагрузки в треугольник, возможны также 6 сочетаний открытых и закрытых состояний транзисторов.
Действующее значение фазного напряжения 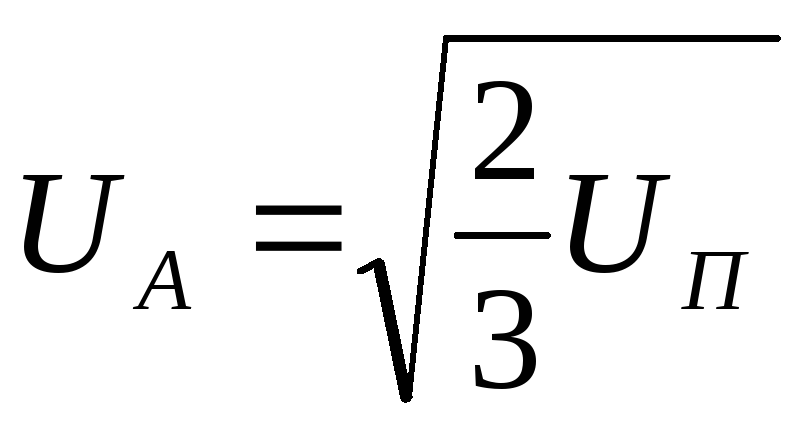
При R-Lнагрузке форма напряжения не изменяется. В выходном напряжении трехфазного АИН при λ=1800отсутствуют четные гармоники и гармоники, кратные трем, а присутствуют: 1, 5, 7, 11…
При соединении нагрузки в звезду фазное напряжение на нагрузке имеет следующий гармонический вид:
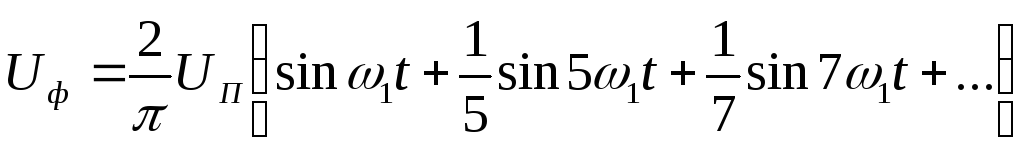 ,
где–
круговая частота 1-ой гармонической
выходного напряжения.
,
где–
круговая частота 1-ой гармонической
выходного напряжения.
Рисунок 39.11 Фазные напряжения нагрузки при соединении нагрузки треугольником, λ=1800
33.Принцип действия трехфазного инвертора напряжения с углом проводимости =180°. Нагрузка активно-индуктивная, 0°< <60°, соединение- звезда.
При активно-индуктивной нагрузке ток отстаёт от напряжения. При этом изменяется конфигурация эквивалентных схем замещения инвертора и в работу вступают возвратные диоды. Но это не приводит к изменению формы напряжения на нагрузке, т.к. в каждый момент времени в схеме остаются включенными три полупроводниковых вентиля.
Конфигурация эквивалентных схем зависит от угла φ(угол сдвига тока относительно напряжения).
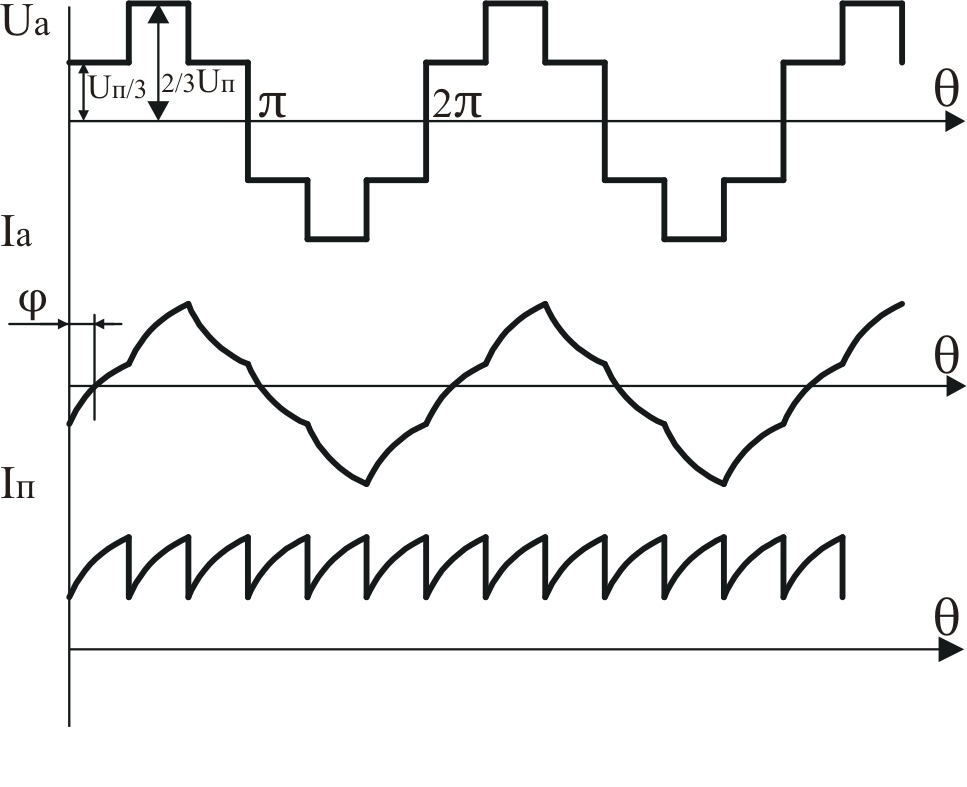
Рисунок 40.1 Временные диаграммы работы трехфазного мостового АИН
с λ=1800 при 0º<φ<60º
В момент времени θ=0отключаетсяVT4и подаётся открывающий импульс наVT1.Однако из-за действия ЭДС самоиндукциифазы А, изменившей свой знак в момент
времениθ=π, открывается диодVD1и токфазы Апротекает по цепифаза
А – VD1 – VT5
– фаза С – фаза А. Т.е. на интервале0<θ<φпроисходит обмен реактивной
энергией междуфазами А и С. При
этом подключение фаз к источнику питания
остаётся аналогичным, как и при активной
нагрузке. Поэтому форма напряжения на
нагрузке не изменяется. В момент времениθ=φ, токI становится равным нулю,VD1закрывается, открываетсяVT1.
Таким образом, приφ<60ºвозврата
энергии в источник питания не происходит
(о чем свидетельствует однополярная
форма токаiП),
а на отдельных интервалах происходит
обмен реактивной энергией между фазами.
Такие же процессы происходят и в других
фазах
становится равным нулю,VD1закрывается, открываетсяVT1.
Таким образом, приφ<60ºвозврата
энергии в источник питания не происходит
(о чем свидетельствует однополярная
форма токаiП),
а на отдельных интервалах происходит
обмен реактивной энергией между фазами.
Такие же процессы происходят и в других
фазах
34.Принцип действия трехфазного инвертора напряжения с углом проводимости =180°. Нагрузка активно-индуктивная, 60°<<1200, соединение- звезда.

Рисунок 41.1 Временные диаграммы работы трехфазного мостового АИН
с λ=1800 при 60º<φ<120º
На интервале времени 0<θ<π/3открытыVT5,VT6иVD1.
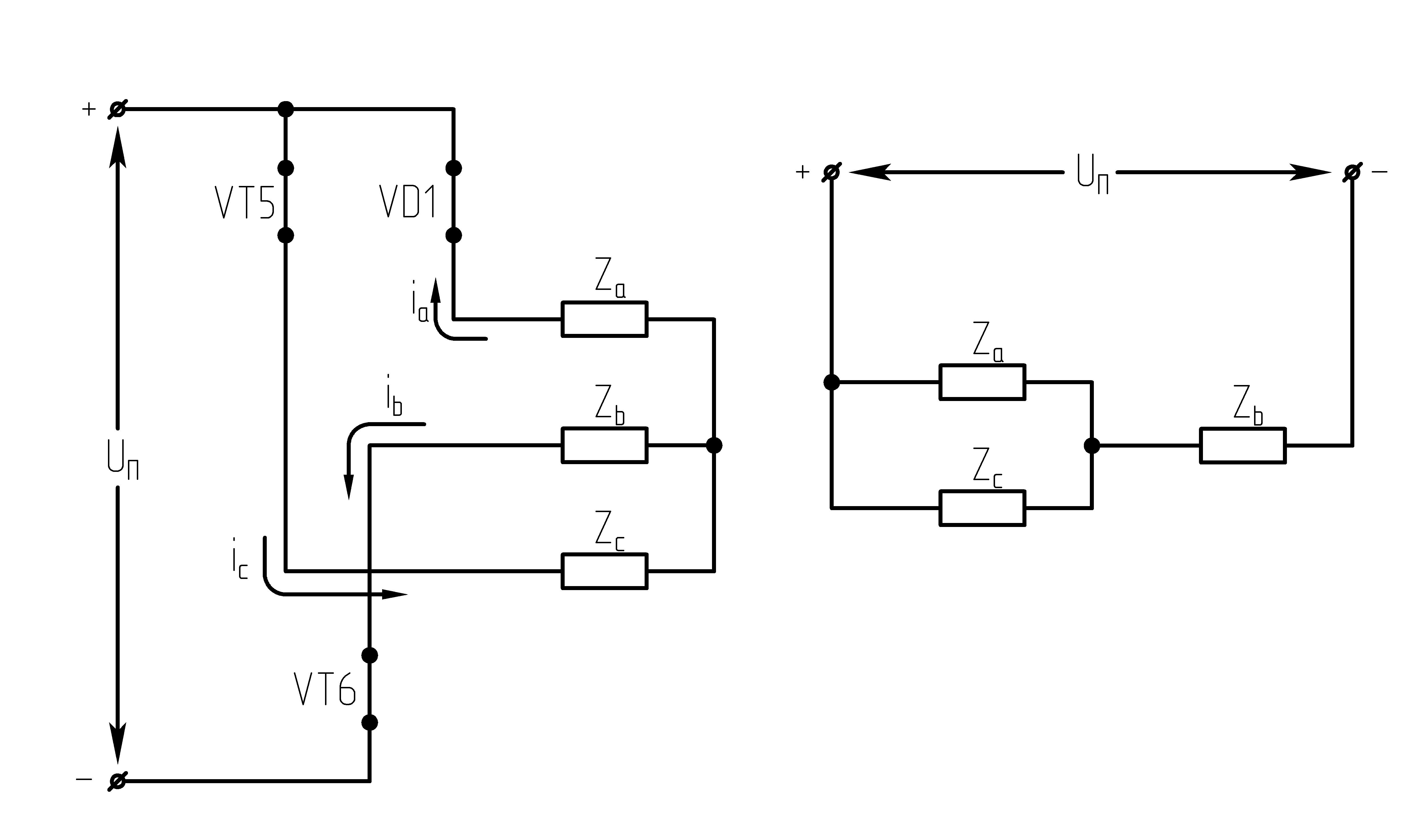
Рисунок 41.2 Эквивалентная схема замещения трехфазного мостового АИН
на интервале 0<θ<π/3
На этом интервале происходит обмен реактивной энергией между фазами А и С. В момент времениθ=π/3закрываетсяVT5и должен открытьсяVT2, но из-за действия ЭДС самоиндукциифазы С, открываетсяVD2.На интерваеπ/3<θ<φVT1 ещё не открылся, а открытыVD1,VT6иVD2(открывается под действием ЭДС самоиндукциифазы С) и часть энергии, запасённой в полях
индуктивностей фаз А и Свозвращается в источник питания, а часть рассеивается в активных сопротивленияхфаз А и С.
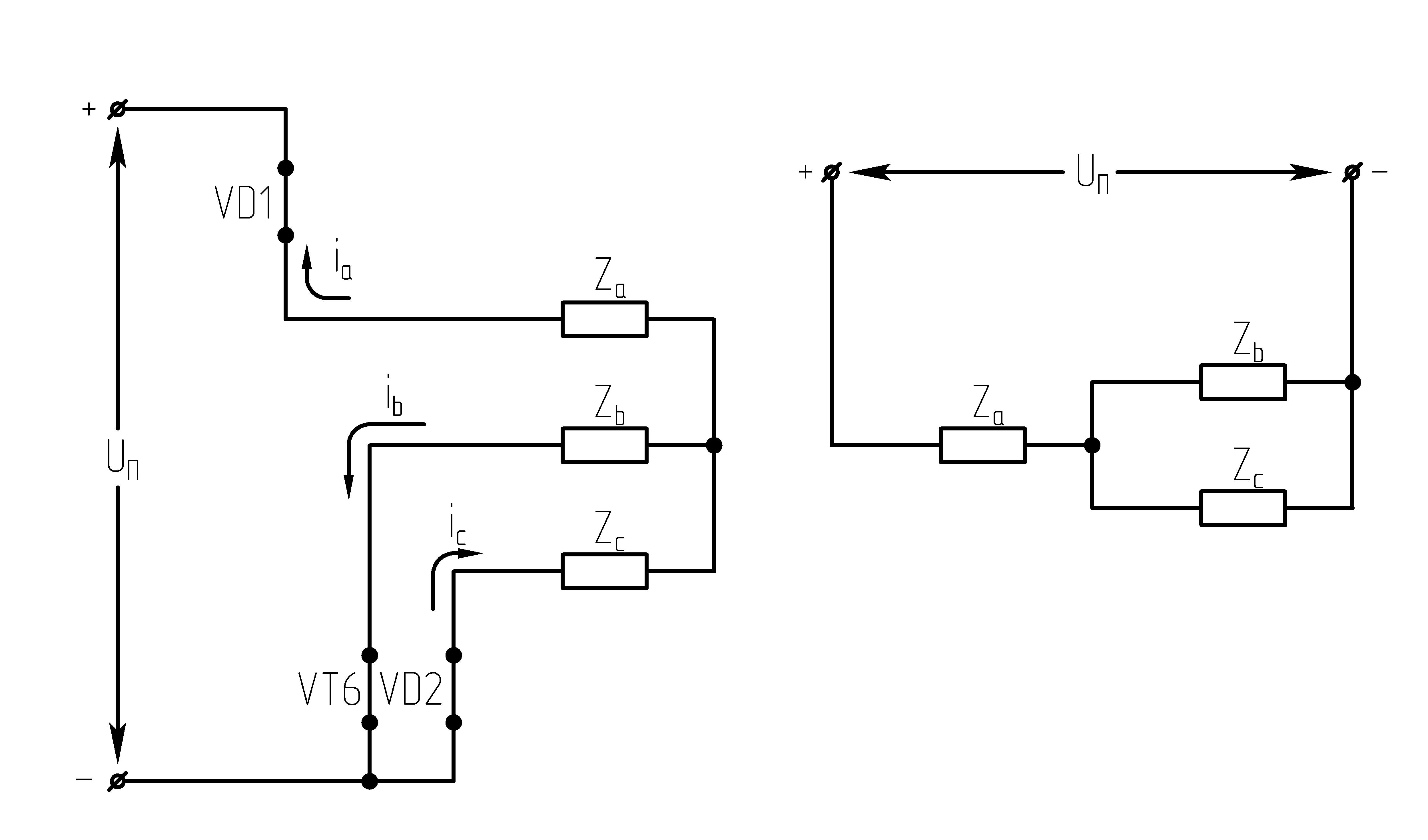
Рисунок 41.3 Эквивалентная схема замещения трехфазного мостового АИН
на интервале π/3<θ<φ
Энергия фазы ВчерезVT6,VD2ифазу Срассеивается в активных сопротивленияхфаз СиВ. На этом интервале токiПменяет свой знак.
Таким образом, можно сделать следующие выводы:
при φ<60ºвозврата энергии в источник питания не происходит (о чем свидетельствует однополярная форма токаiП), а на отдельных интервалах происходит обмен реактивной энергией между фазами.
при φ>60ºчасть энергии, запасённой в полях индуктивностейфаз, возвращается в источник питания, а часть рассеивается в активных сопротивленияхфаз .
Рисунок 47.2 Временные диаграммы работы инвертора при φ<π/3 и φ>π/3
35.Принцип действия трехфазного инвертора напряжения с углом проводимости =120°. Нагрузка активная, соединение- звезда.
Рисунок 42.1 Схема трехфазного АИН
Алгоритм переключения ключей инвертора при λ=1200выглядит следующим образом:
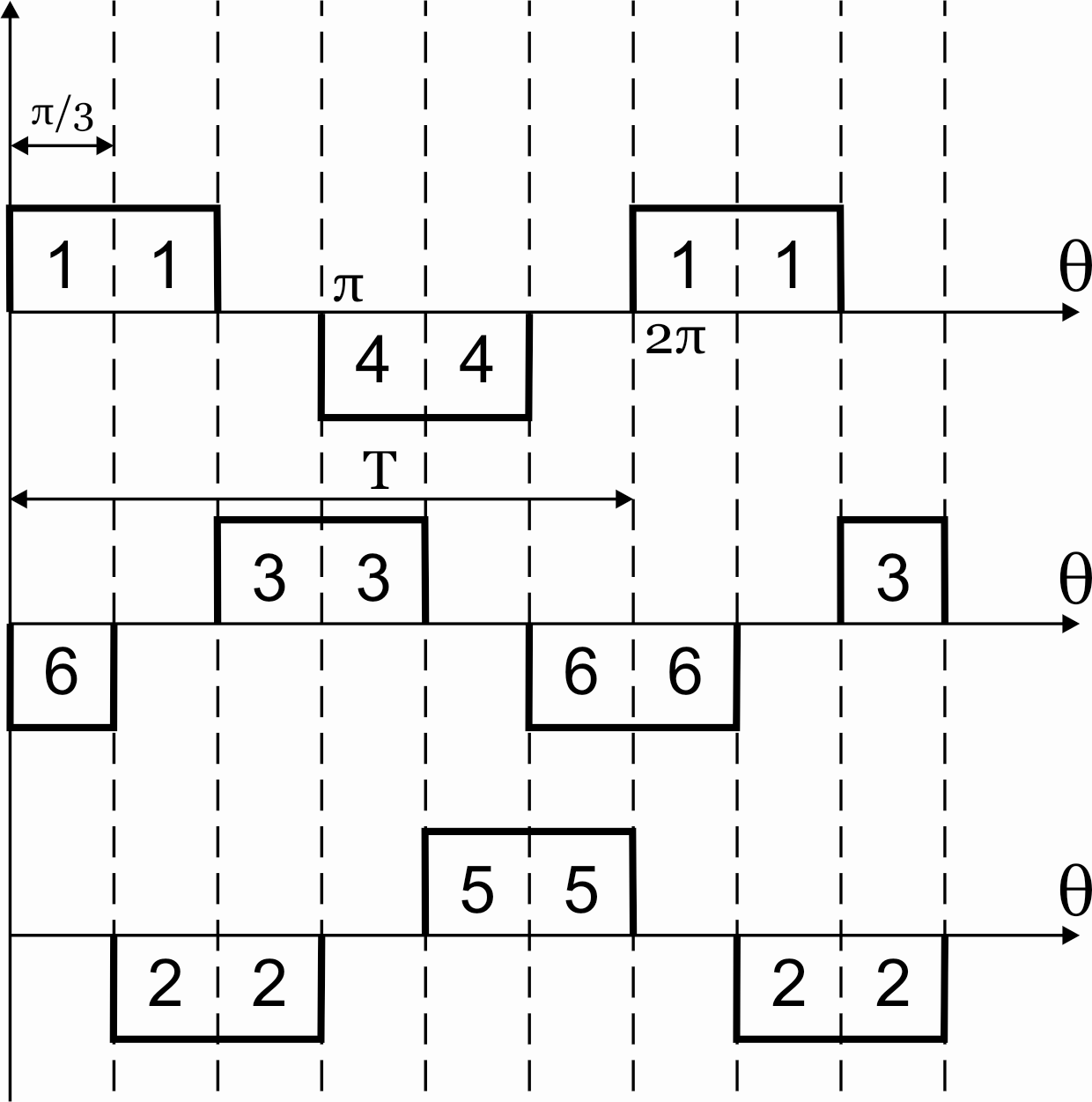
Рисунок 42.2 Алгоритм переключения ключей АИН при λ=1200
Как видно из алгоритма переключения ключей инвертора при λ=1200в схеме в каждый момент времени открывающие импульсы одновременно подаются лишь на два тиристора. При этом фазы нагрузки соединены между собой последовательно, а одна из них отключена (Uф=0). Следовательно, к соответствующим работающим фазам прикладывается напряжениеUф=± UП /2.За периодТвыходного напряжения возможны 6 независимых сочетаний открытых и закрытых состояний вентилей.
 ;
;- ;
- ;
- ;
- ;
- .
 ;
; ;
;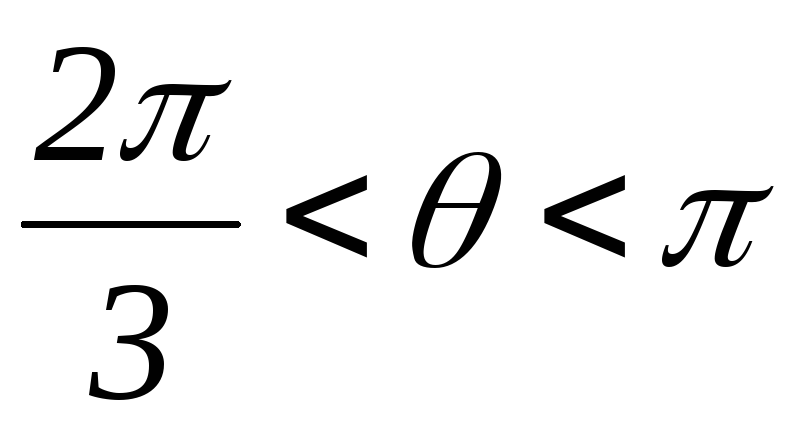 ;
; ;
;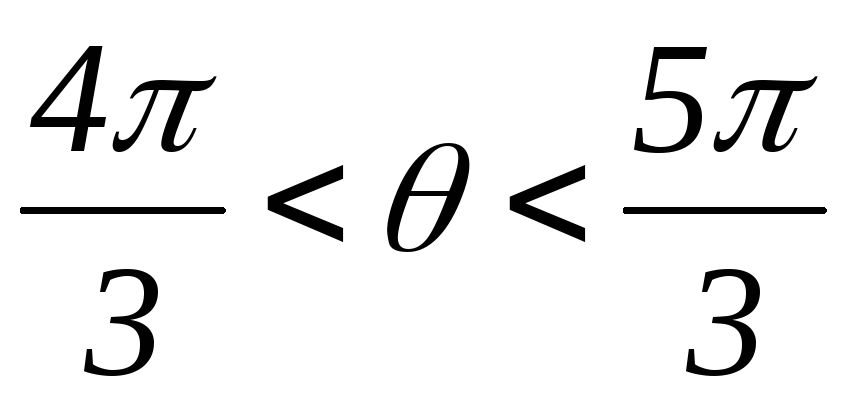 ;
;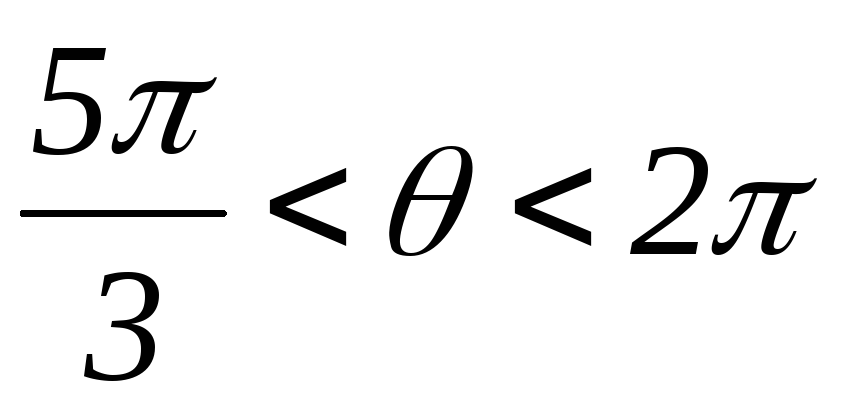 .
. 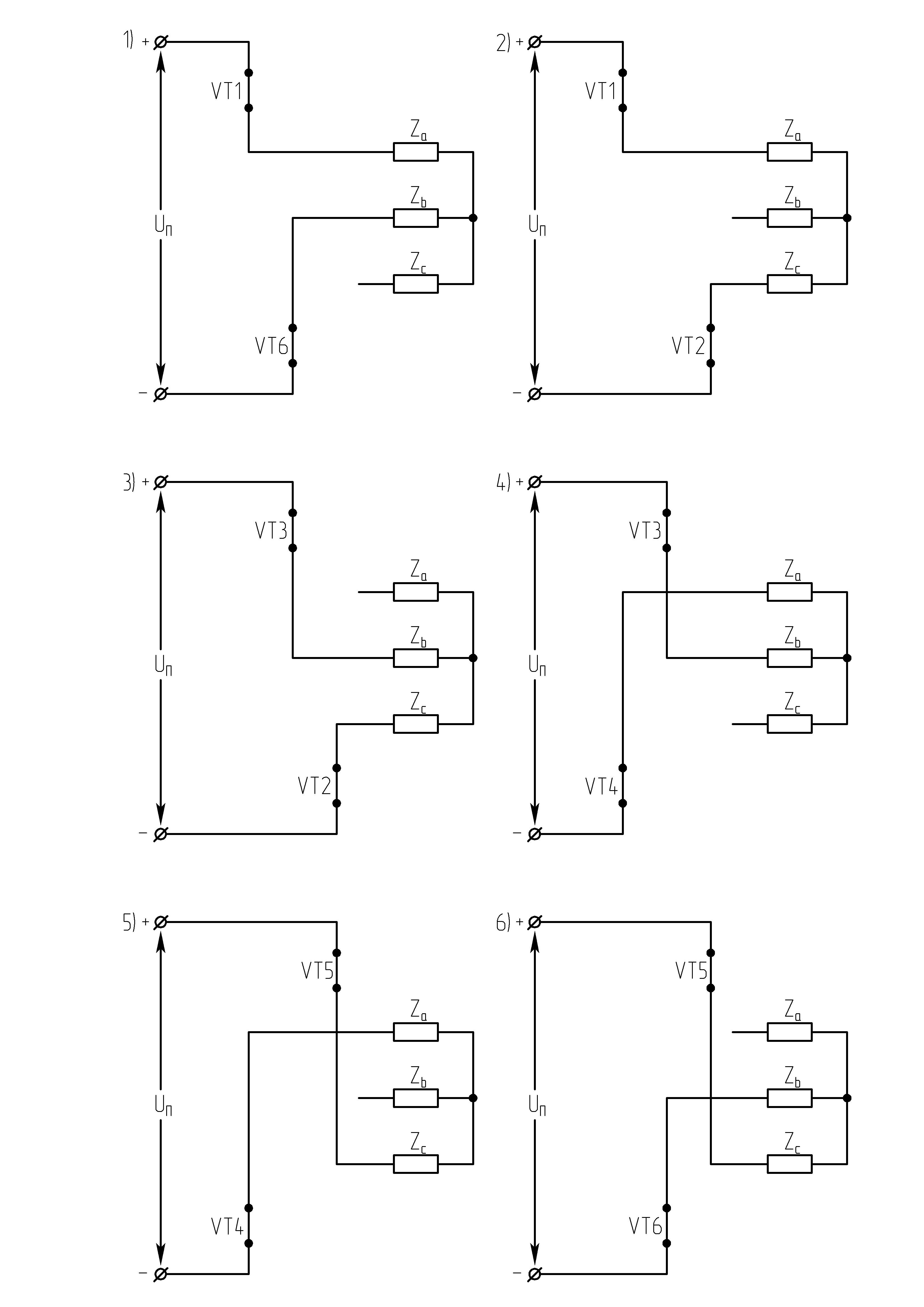
Рисунок 42.3 Схемы замещения АИН при λ=1200 для различных
временных интервалов
Таким образом, фазы подключены между собой последовательно, т.е. к ним прикладываются напряжения ± UП /2.
Вид фазных напряжений показан на рисунке 42.4.
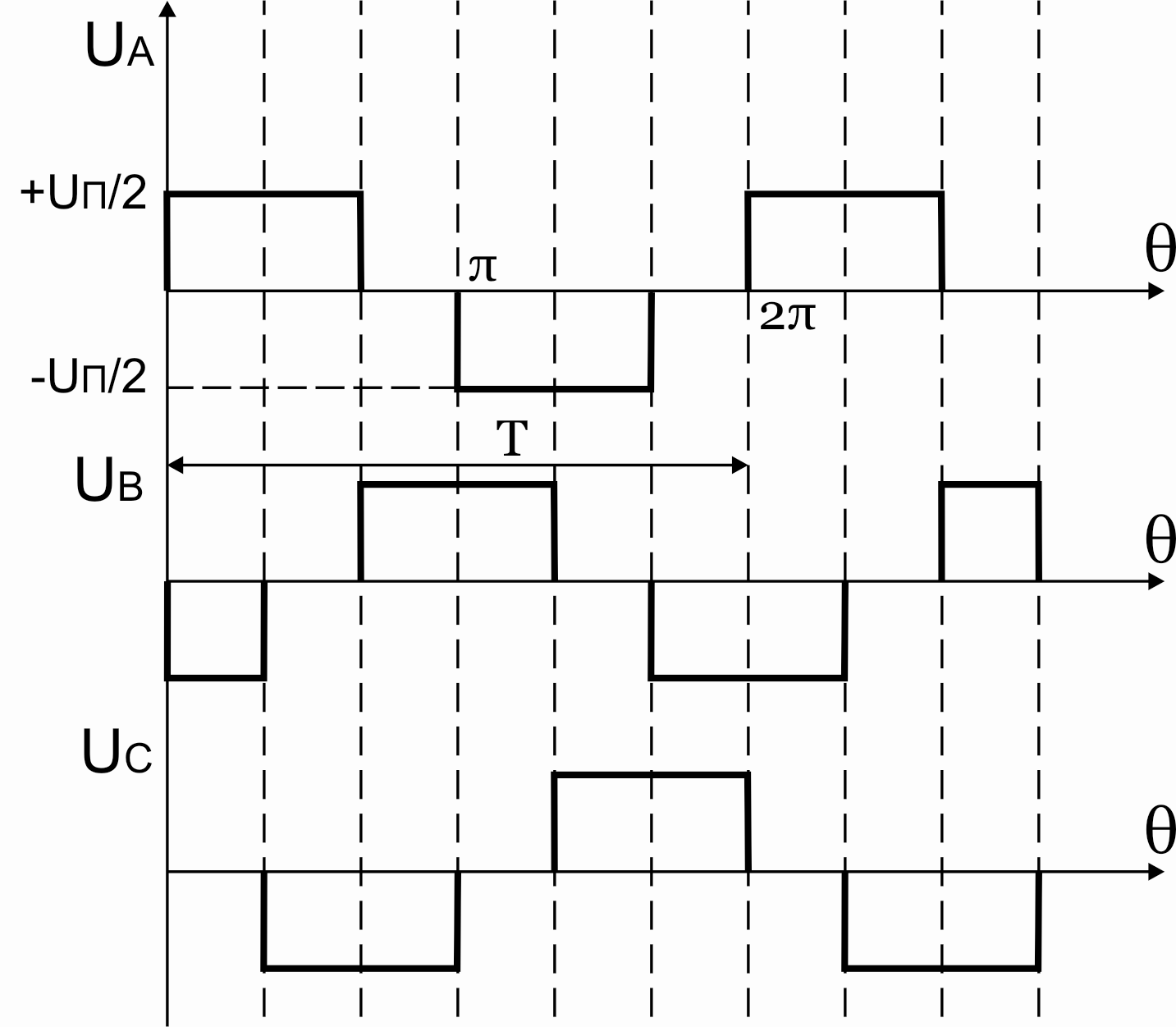
Рисунок 42.4 Фазные напряжения АИН при λ=1200 и активной нагрузке
36.Принцип действия трехфазного инвертора напряжения с углом проводимости =150°. Нагрузка активная, соединение- звезда.
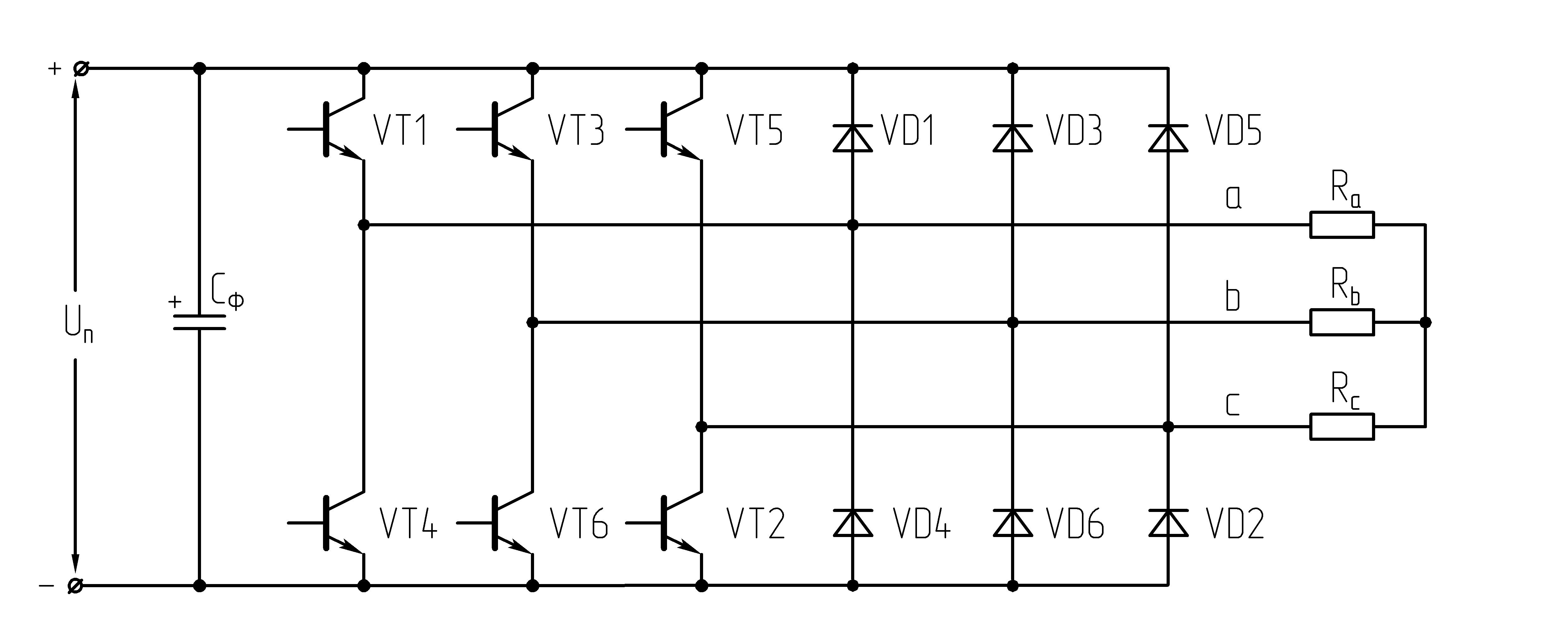
Рисунок 44.1 Схема трехфазного мостового АИН
При λ=1500алгоритм
переключения ключей инвертора следующий:
Рисунок 44.2 Алгоритм переключения ключей инвертора
Выполнение данного алгоритма позволяет получить на нагрузке кривую трехфазного симметричного несинусоидального переменного напряжения.
Из алгоритма переключения инвертора
следует, что в трехфазном мостовом АИН
при λ=1200за периодТвыходного напряжения инвертора имеется
12 независимых сочетаний открытых и
закрытых состояний транзисторов. При
этом , в каждый момент времени открыты
либо три, либо два ключа.1.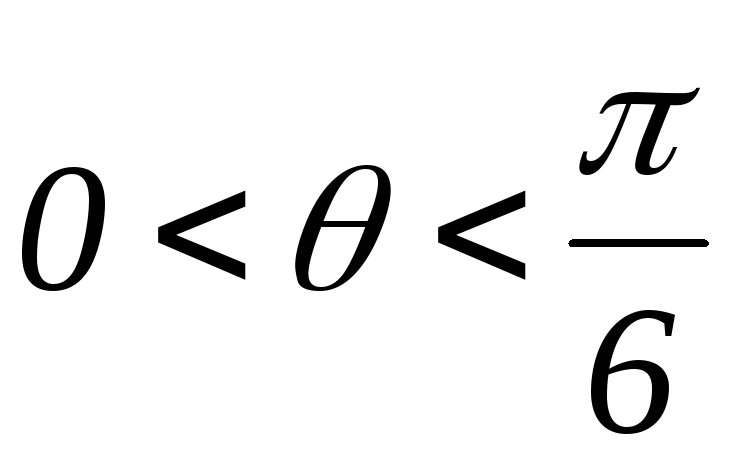
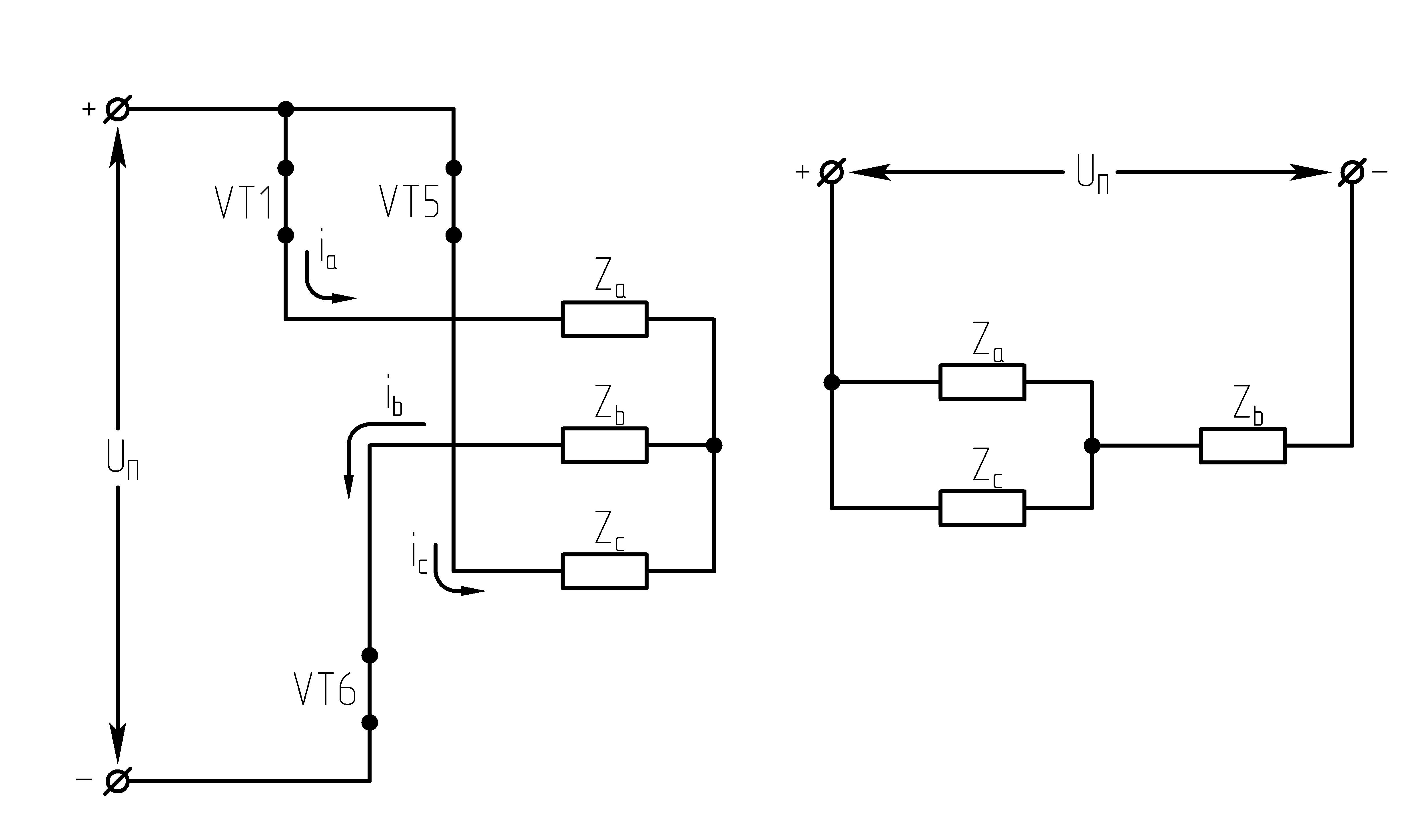 На
данном интервале открытыVT1,VT6,VT5.
Схема замещения следующая:
На
данном интервале открытыVT1,VT6,VT5.
Схема замещения следующая:
Рисунок 44.3 Схема замещения при 0º<φ<30º
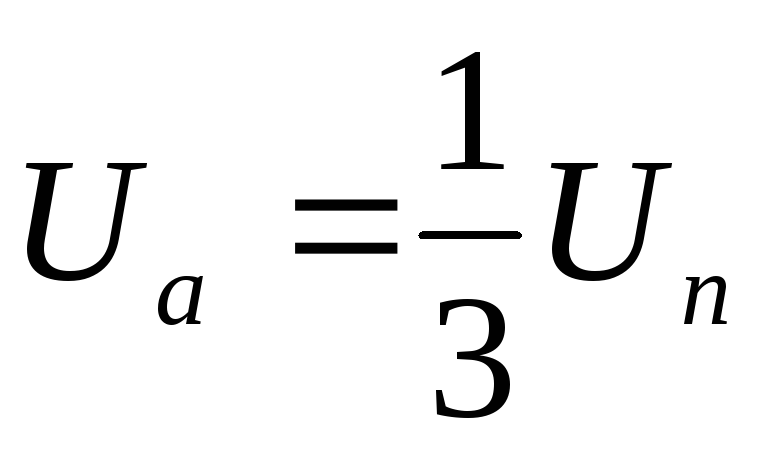 2.
2.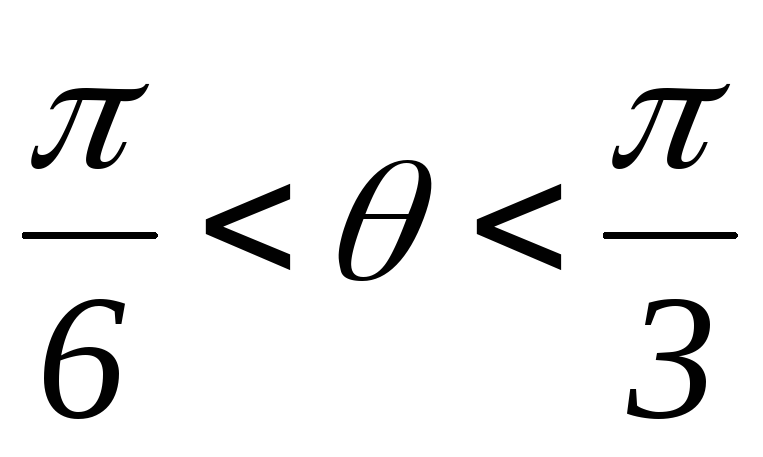 На
данном интервале открытыVT1иVT6. Схема замещения
следующая:
На
данном интервале открытыVT1иVT6. Схема замещения
следующая:
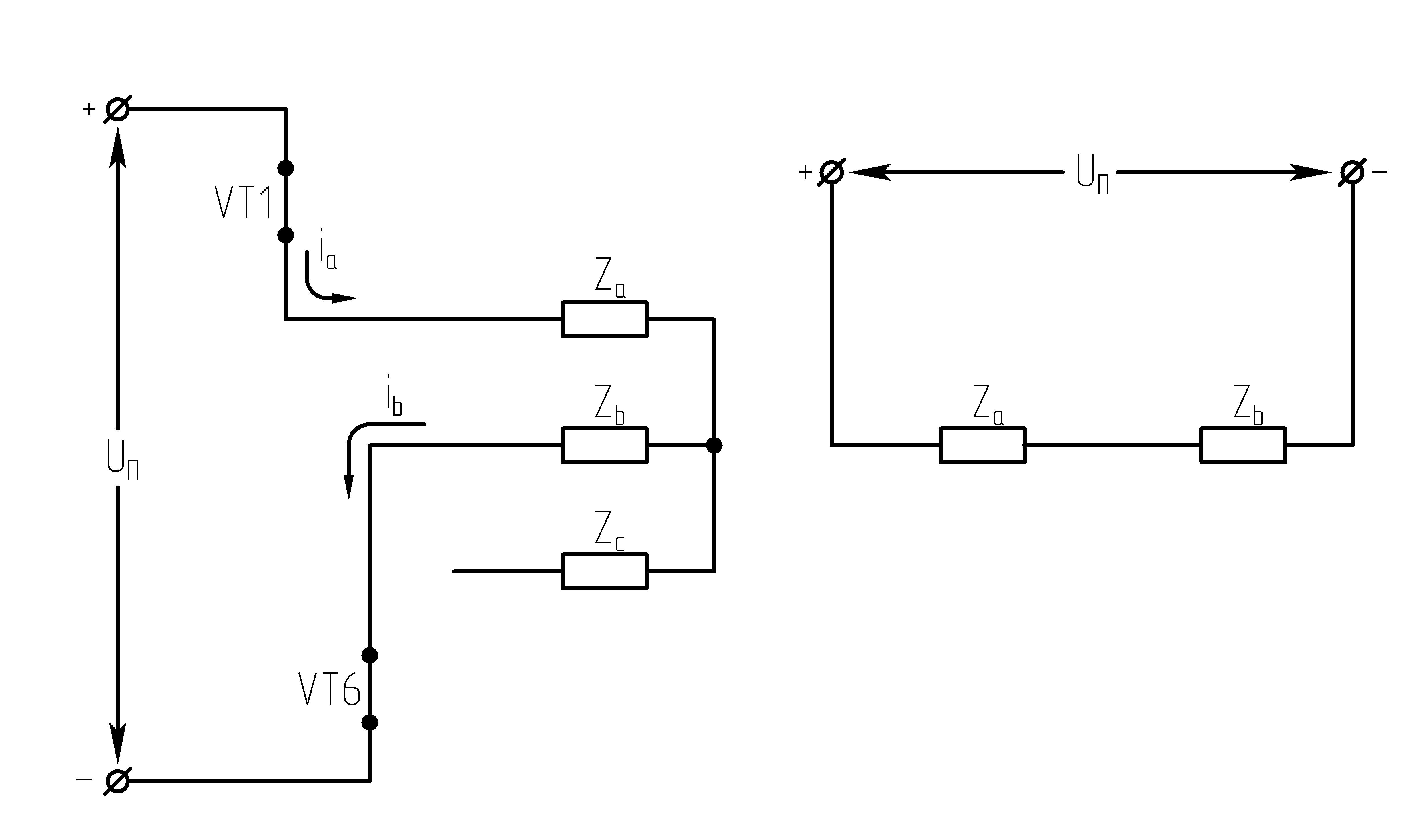
Рисунок 44.4 Схема замещения при 30º<φ<60º
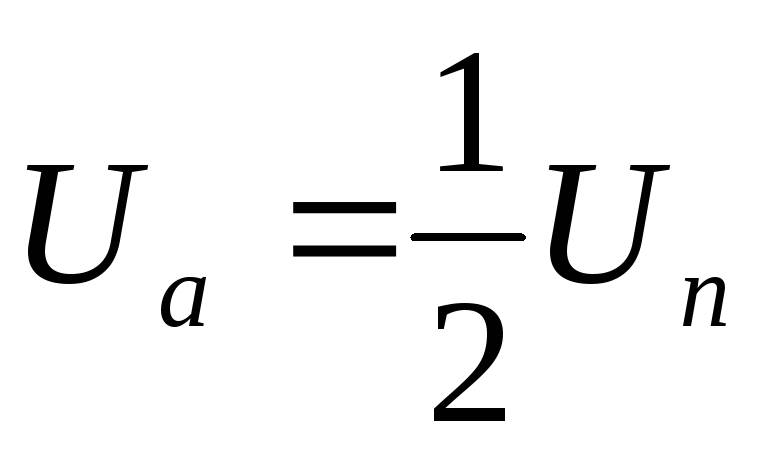
3. 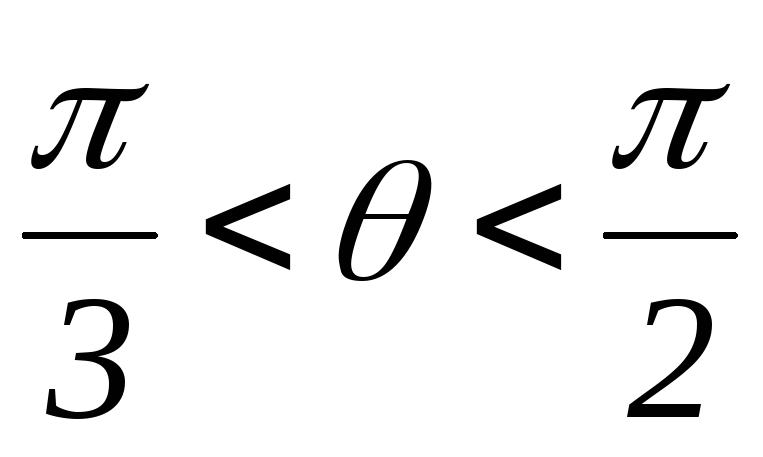
На данном интервале открыты VT1,VT2,VT6. Схема замещения следующая:
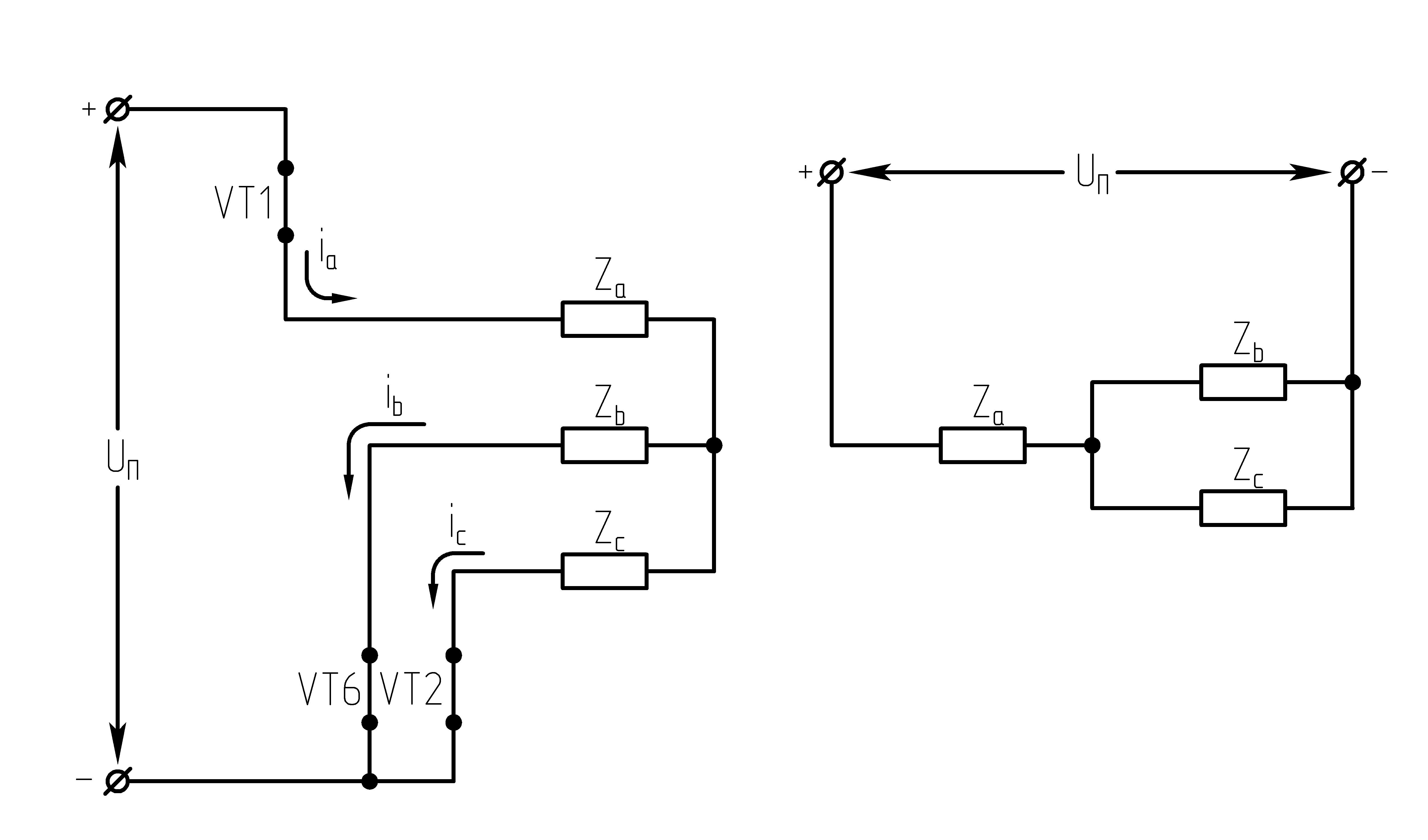
Рисунок 44.5 Схема замещения при 60º<φ<90º
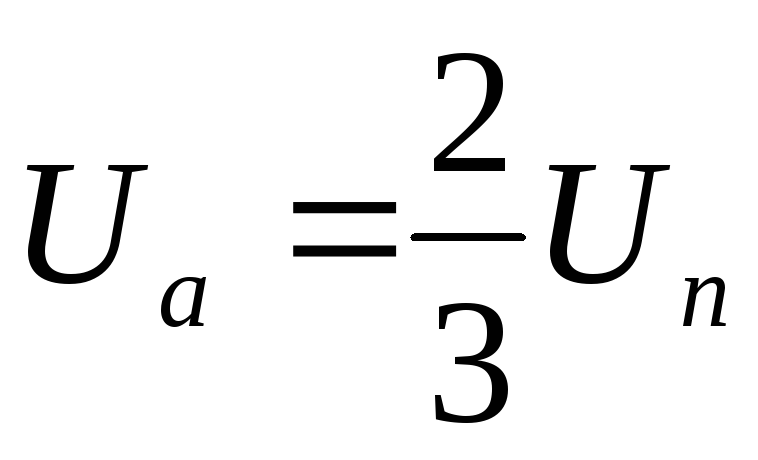
4. 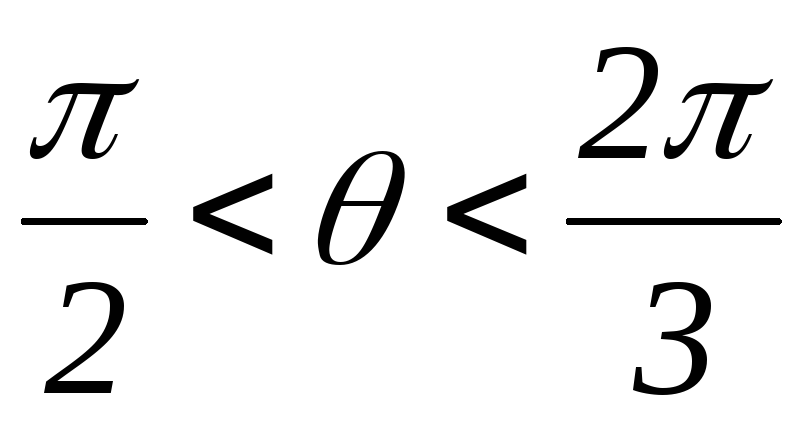
На данном интервале открыты VT1иVT2. Схема замещения следующая:
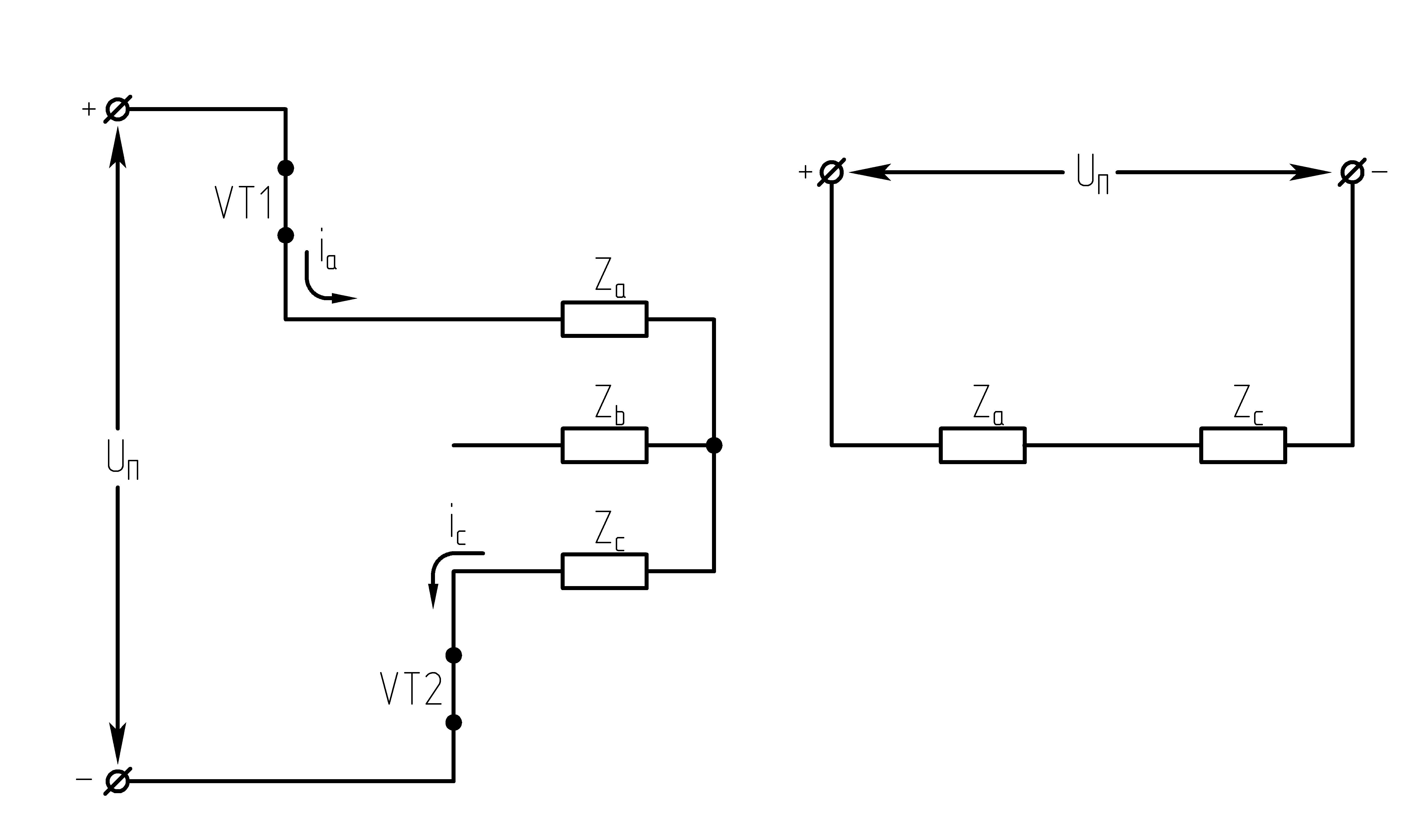
Рисунок 44.6 Схема замещения при 90º<φ<120º
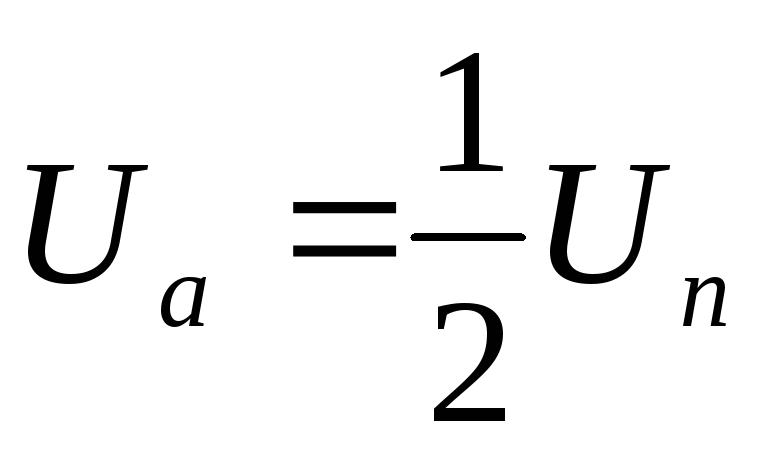
5.  На
данном интервале открытыVT1,VT3,VT2.
Схема замещения следующая:
На
данном интервале открытыVT1,VT3,VT2.
Схема замещения следующая:

Рисунок 44.7 Схема замещения при 120º<φ<150º

6.  На
данном интервале открытыVT3иVT2. Схема замещения
следующая :
На
данном интервале открытыVT3иVT2. Схема замещения
следующая :

Рисунок 44.8 Схема замещения при 150º<φ<180º
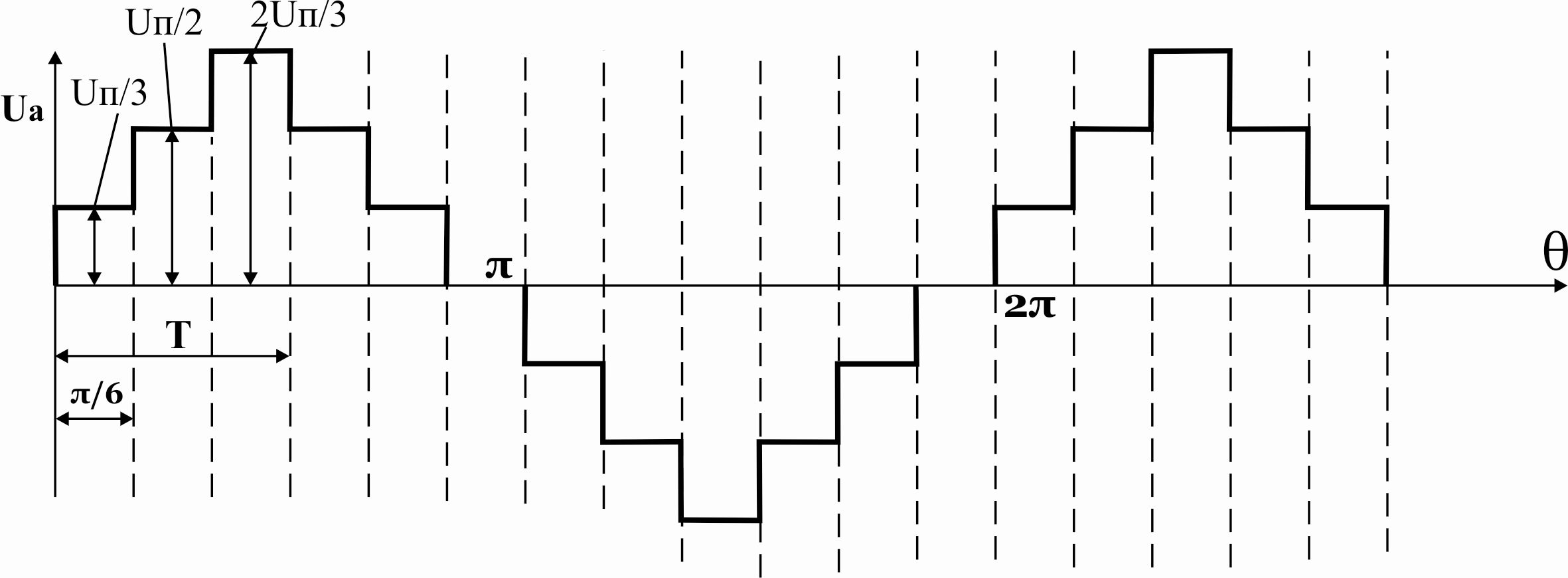
Рисунок 44.9 Напряжение фазы А трехфазного мостового АИН
при λ=1500 и активной нагрузке
Из схем замещения инвертора видно, что к каждой фазе нагрузки за
период Твыходного напряжения прикладываются напряжения: