


Провод пнсв: особенности и характеристики
Провод типа ПНСВ
Провод для прогрева бетона ПНСВ получил широкое распространение в строительстве. В отличие от других проводов его основное назначение не передача электрической энергии, а нагрев участков бетона в которые он укладывается. Преимущественно его используют во время строительства в зимнее время, но иногда его применяют в качестве системы теплых полов. Но давайте обо всем по порядку.
Расшифровка и конструкция провода ПНСВ
Начать наше знакомство с проводом ПНСВ предлагаем с расшифровки его названия, которое уже многое скажет о его назначении. Более же дательный разбор конструкции провода позволит нам определиться с основными его характеристиками.
Расшифровка названия провода ПНСВ
Маркировка провода ПНСВ несколько отличается от аббревиатур обычных проводов. В связи с этим у многих возникают определенные трудности.
Провода ПНСВ
Итак:
- Первая буква «П» говорит нам о том, что перед нами провод.

- Вторая буква «Н» и она является наиболее определяющей в назначении кабеля. Именно она указывает на то, что провод нагревательный и использовать его для передачи электрической энергии не стоит.
- Третья буква «С». Она указывает нам на материал, из которого изготовлен проводник. В нашем случае провод имеет стальной проводник. А как известно сталь имеет не самый высокий показатель теплопроводности, что и позволяет применять провод в качестве нагревательного.

Обратите внимание! Для изготовления провода может применяться сталь двух видов – оцинкованная или не оцинкованная. Первый вариант позволяет защитить провод от коррозии, но как вы понимаете цена такого изделия несколько выше. Хотя разница в цене не столь существенна.
Конструкция провода ПНСВ
- Последняя буква аббревиатуры «В». Она указывает на материал, из которого изготовлена изоляция провода. В данном случае это винил или как его правильнее называть поливинилхлорид.
 В данном случае это винил или как его правильнее называть поливинилхлорид.
В данном случае это винил или как его правильнее называть поливинилхлорид.После этого обычно указывается сечение провода в мм2. Данная марка провода имеет не столь широкий модельный ряд. Наиболее распространенными моделями являются изделия в 1, 1,2, 1,4 мм
Конструкция провода ПНСВ
Провод прогревочный ПНСВ по своей конструкции очень похож на провод ПВ1, который по сути является его прототипом. Тем не менее давайте уделим несколько слов его конструкции.
- Прежде всего это конечно жила провода. Она выполняете одной цельной проволокой, что конечно же оказывает серьезное влияние на общую гибкость провода. Особенно это заметно в проводах большего сечения. По классификации гибкости данный провод относят к первому наименее гибкому классу.
Гибкость проводов ПНСВ
- В то же время характеристики гибкости данный провод имеет относительно неплохие. Так минимальный радиус изгиба должен составлять не менее 5 наружных диаметров провода, что по характеристике соизмеримо с проводами 3-го и более высоких классов гибкости проводов.
- Кроме непосредственно жилы наш греющий провод имеет изоляцию. Она напрямую зависит от сечения провода. Ведь чем больше сечение, тем толще должна быть изоляция. Так минимальная толщина ПВХ-изоляции составляет 0,8 мм. Но для изделий сечением до 4 мм2 она может составлять 1 мм.
Характеристики проводов ПНСВ
- Что касается расцветки провода, то обычно к этому не предъявляют особых требований. Стандартной является черная или коричневая окраска. Но по желанию заказчика возможен выпуск проводов и другой окраски.
Характеристики провода ПНСВ
Характеристики на ПНСВ провод прогревочный во многом отличаются от характеристик обычных проводов. Ведь к нему предъявляются совершенно другие требования, в данном типе провода на первое место выходят не свойства проводника и изоляции, а температурные характеристики и теплоотдача.
- Если говорить о температурных характеристиках, то для провода ПНСВ часто указывают максимально допустимую температуру, которая равна +80⁰С. Но это та температура выше которой уже происходит разрушение изоляции. А вот во время эксплуатации инструкция советует соблюдать температуру в пределах – 60⁰С — +50⁰С. То есть нагрев провода выше +50⁰С не рекомендуется.
 Но это та температура выше которой уже происходит разрушение изоляции. А вот во время эксплуатации инструкция советует соблюдать температуру в пределах – 60⁰С — +50⁰С. То есть нагрев провода выше +50⁰С не рекомендуется.
Но это та температура выше которой уже происходит разрушение изоляции. А вот во время эксплуатации инструкция советует соблюдать температуру в пределах – 60⁰С — +50⁰С. То есть нагрев провода выше +50⁰С не рекомендуется.Монтаж проводов ПНСВ при низких температурах
- Еще одной важной деталью является температура монтажа. Хотя эксплуатация провода допускается при температуре до — 60⁰С, но его монтаж не стоит производить при температуре ниже — 15⁰С.
- Следующей важной характеристикой у данного типа провода является удельная мощность тепловыделения. У обычных проводов данный параметр не превышает 1 – 3%, но нам необходимо дабы этот параметр был как можно выше. Обычно производители заявляют удельную мощность в районе 20Вт/м.
Обратите внимание! Некоторые производители заявляют удельную мощность до 40Вт / м, но здесь многое зависит от температуры для которой производился расчет и поверхности. Так поверхности с армированием позволяют увеличить данный показатель.

- Еще одним важным параметром является удельное сопротивление провода. Оно напрямую зависит от сечения. Так ПНСВ 1,2 провод имеет сопротивление равное 0,12Ом/м, а изделие сечением в 2 мм2 имеет сопротивление равное 0,044Ом/м.
Основные характеристики проводов НПСВ
Подключение провода ПНСВ
Если вы собрались выполнять монтаж провода ПНСВ своими руками, то приведем вам основные правила монтажа. Ведь для получения требуемых показателей его следует выполнять не на глаз, а применять специальный расчет.
В этом расчете должны учитываться площадь предполагаемых работ, объем бетона, скорость ветра на месте проведения работ, температурные показатели, требуемое время прогрева бетона и даже схему подключения провода.
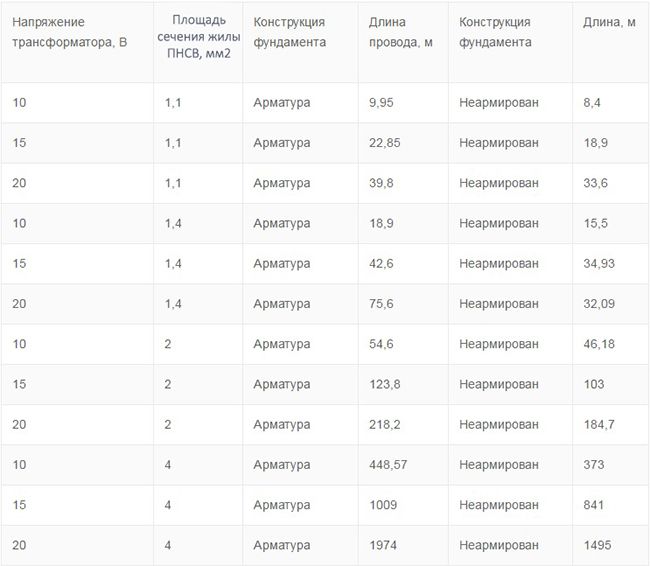
Расчет длины проводов ПНСВ для прогрева | На самой первой стадии выполняется расчет требуемого количества провода. Он зависит от объема бетона, схемы подключения и наружных температур. Подключать весь провод одним куском нельзя, так как это не даст должно эффекта. Обычно подключение выполняется отдельными секциями длиной 17 или 28 метров каждый. |
Схемы трехфазного подключения | Подключение может быть выполнено двухфазным или трехфазным. При это трехфазной подключение может быть выполнено по схеме звезды или по схеме треугольника. Как вы можете видеть на видео выбор схемы подключения производится исходя из местных условий. |
Нагрузка отдельных секций НПСВ | Еще одним важным условием выбора длины провода является соблюдение токовых нагрузок. Так на каждом участке провода сила тока должна быть приблизительно равна 15А.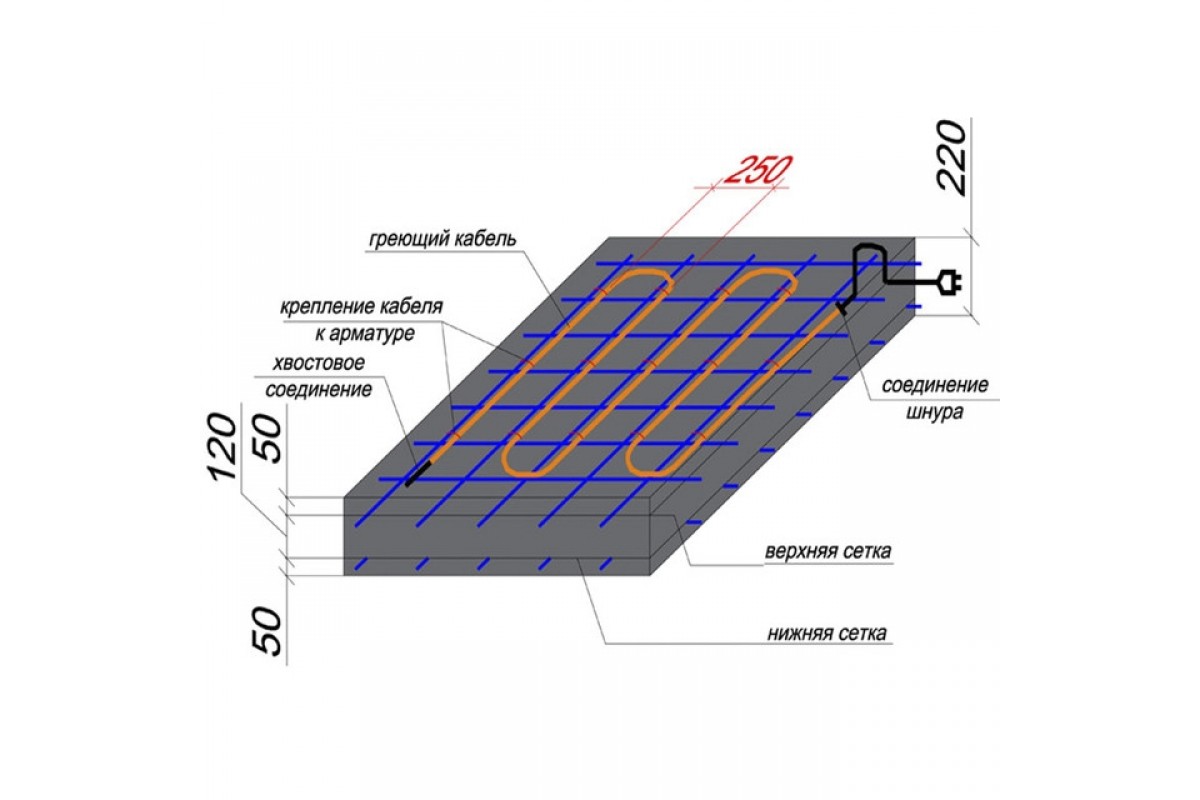 |
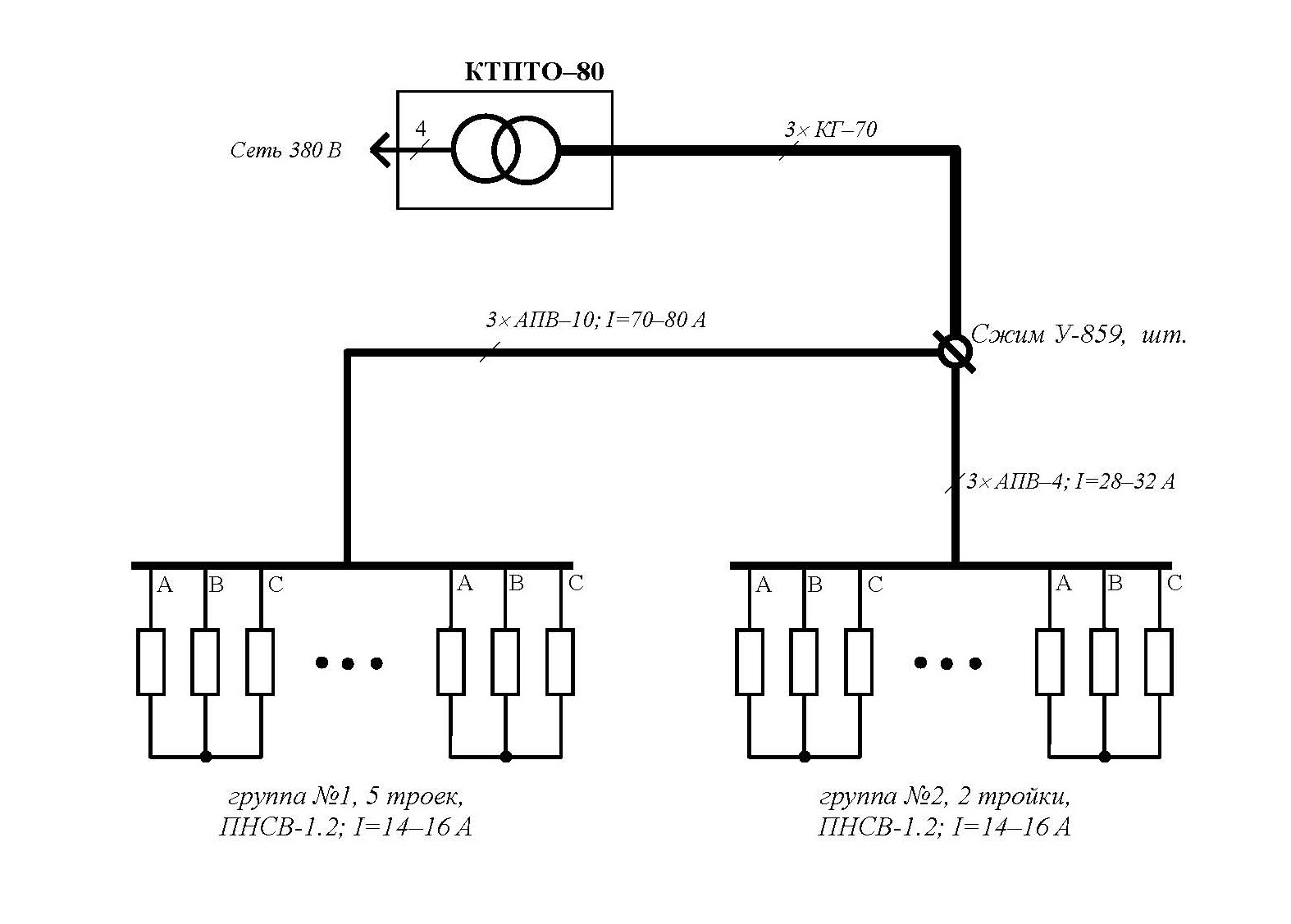
Схема подключения нагревающего провода | Чтоб ток ПНСВ и температура провода соответствовала требованиям обычно на него подают напряжение в 70 – 100В. Для этого используют специальный понижающий трансформатор. Одной из наиболее распространенных моделей для этого является трансформатор КТПТО-80. |
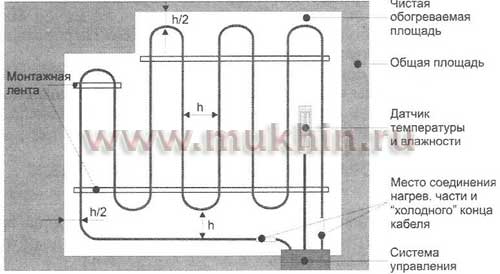
Правила монтажа проводов ПНСВ | Теперь можно перейти непосредственно к правилам укладки провода. Он должен укладываться равномерно по всей обогреваемой площади. Между отдельными проводами следует соблюдать расстояние не менее 5 см. При этом сам провод в любом случае не должен соприкасаться между собой. Соприкосновение с арматурой не рекомендуется, но обычно этим правилом пренебрегают. Для улучшения прогрева бетона кабель рекомендуется укрывать слоем фольги. Ее толщина должна быть в пределах 2 – 2.5 мм. |
На фото структурная схема подключения проводов НПСВ | Подключение провода ПНСВ к трансформатору должно выполняться обычным проводом. Зачастую для этого используют провод ПВ1, который как мы знаем является прототипом нашего греющего провода. Зачастую для этого используют провод ПВ1, который как мы знаем является прототипом нашего греющего провода. |
Обратите внимание! Провода ПНСВ и ПВ1 или любой другой обычный провод должны соединяться непосредственно в бетоне. Особых требований к качеству соединения не предъявляют, это должно быть любое качественное соединение, соответствующее правилам ПУЭ и способное обеспечить должную влагозащищенность.
Вывод
ПНСВ провод для прогрева бетона, который за вполне приемлемую цену позволяет решать проблемы, связанные с бетонными работами в зимнее время. При этом он не предъявляет особых требований по подключению и эксплуатации.
Главной трудностью является расчет необходимой длины и выбор схемы подключения. Но незначительные ошибки на данной стадии вполне возможно компенсировать за счет изменения рабочего напряжения провода.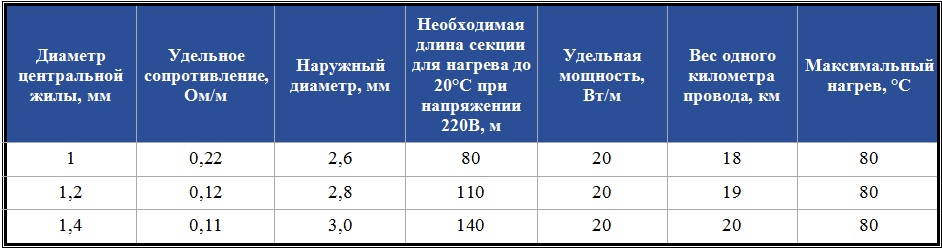 Исходя из всего этого провод типа ПНСВ вполне возможно рекомендовать для применения.
Исходя из всего этого провод типа ПНСВ вполне возможно рекомендовать для применения.
Провод ПНСВ. Устройство и применение. Маркировка и параметры
Чтобы прогреть бетон в холодное время года при проведении строительных работ используют специальную марку провод ПНСВ. Если его сравнивать с другими видами проводников, то он служит не для транспортировки электроэнергии, а для нагрева тех участков бетона, куда он уложен. Больше всего его применяют при строительстве в морозную погоду, однако существует и другое применение этому проводу, например, в теплых полах.
Как устроен провод ПНСВ
Его устройство похоже на провод ПВ1, являющегося его аналогом. Но эту марку следует рассмотреть отдельно.
- Токоведущая жила изготавливается из стали, одной цельной проволокой. Это значительно снижает ее гибкость, что особенно заметно на толстых проводах. Провод ПНСВ относится к 1-му классу гибкости по принятой классификации. Однако на самом деле его гибкость значительно выше. Наименьший радиус изгиба завод изготовитель устанавливает 5 внешних диаметров, что по параметрам соответствует проводникам более высоких классов.
- Греющий провод снаружи покрыт изоляционным слоем поливинилхлорида, толщина которого зависит от сечения жилы. Чем толще токоведущая жила, тем больше слой изоляции. Минимальная толщина изоляции ПВХ составляет 0,8 мм.
- Цветовая гамма изоляции довольно широкая, так как к цвету особые требования по нормативным документам не предъявляются. Чаще всего оболочку изготавливают коричневого или черного цвета, хотя можно заказать любой цвет изоляции.
 Наименьший радиус изгиба завод изготовитель устанавливает 5 внешних диаметров, что по параметрам соответствует проводникам более высоких классов.
Наименьший радиус изгиба завод изготовитель устанавливает 5 внешних диаметров, что по параметрам соответствует проводникам более высоких классов.Обозначение этого провода имеет отличия от других марок, поэтому расшифровка многим людям непонятна:
- «П» означает, что это провод, а не кабель.
- «Н» определяет основное назначение этого изделия, и указывает, что этот проводник относится к нагревательным видам, и применять его для транспортировки электроэнергии не следует.
- «С» обозначает материал токоведущей жилы. Для этой марки таким материалом является сталь, которая обладает далеко не лучшим параметром теплопроводности, что и дает возможность использовать ее в качестве нагревательного элемента. Для производства токоведущих жил может использоваться две категории стальной проволоки: оцинкованной и не имеющей покрытия. Оцинкованная жила защищена от появления ржавчины, но стоимость таких изделий больше остальных.
- Буква «В» означает, что материалом изоляции проводника является поливинилхлоридная изоляция (ПВХ-оболочка).
- После буквенной маркировки идут цифровые обозначения, которые указывают сечение стальной жилы в квадратных миллиметрах. Диапазон сечений находится в пределах от 1 до 6 мм2.
Характеристики греющего провода значительно отличаются от параметров обычных проводников, так как к нему предъявляются совсем другие требования.
 В этом виде проводов наиболее важными являются не свойства изоляции и токопроводящей жилы, а величина теплоотдачи и температурные свойства.
В этом виде проводов наиболее важными являются не свойства изоляции и токопроводящей жилы, а величина теплоотдачи и температурные свойства.- Если рассматривать температурные параметры, то для провода ПНСВ обычно указывают допускаемую наибольшую температуру, величина которой +80 градусов. При более высокой температуре изоляция начинает разрушаться.
- Эксплуатационная температура для этой марки проводника находится в интервале -60 +50 градусов.
- Другой важной характеристикой является монтажная температура. Работа провода разрешается при морозах до -60 градусов, но его укладка не должна производиться ниже -15 градусов.
- Не менее важным параметром является удельная мощность выделения тепла. У проводников, предназначенных для передачи электричества, эта характеристика очень низкая. Для греющего провода производителя указывают мощность выделения тепла около 20 ватт на метр. Иногда изготовители заявляют этот параметр величиной до 40 ватт на метр, но здесь играет большую роль расчетная температура, а также наличие армирования поверхности, что значительно повышает этот параметр.

Следующим параметром греющего провода является его удельное сопротивление, которое прямо зависит от сечения токоведущей жилы. Например, ПНСВ сечением 1,2 кв. мм обладает удельным сопротивлением 0,12 Ом на метр, а провод 2 кв. мм – 0,044 Ом на метр.
- Если учесть, что этот вид проводника укладывается в бетонном растворе, то важным его свойством является устойчивость к проникновению воды, а также соляной и кислотной среде, что особенно важно для растворов, которые заливают при отрицательных температурах. В такой бетон обычно добавляют разные присадки для достижения необходимой вязкости.
Для самостоятельного выполнения монтажа греющего провода необходимо соблюдать определенные правила, так как для получения хорошего результата монтаж следует делать не на «глазок», а путем точных расчетов. При этом следует учесть объем бетона, площадь работ, температурный режим, скорость ветра в районе производства бетонных работ, необходимое время прогревания бетона, а также электрическую схему подключения.
- В самом начале производится расчет необходимого количества провода, зависящего от рассмотренных выше параметров.
- Провод ПНСВ следует подключать отдельными секциями. Подключение одним куском нецелесообразно, так как это не дает ожидаемого эффекта.
- Вид подключения может быть трехфазным или двухфазным, а также по схемам треугольника или звезды.
- Важной особенностью этой работы является подбор длины провода и соблюдение нагрузки по току. На отдельной секции сила тока не должна превышать 15 ампер.
- Другой особенностью подключения является соответствие подаваемого напряжения температуре провода. Чаще всего подключают питание в пределах 70-100 вольт, применяя для этого понижающий трансформатор. Наиболее популярной моделью трансформатора при проведении бетонных работ с использованием греющего провода стала марка КТПТО – 80.
- Провод ПНСВ должен равномерно укладываться по всей площади обогрева.
- Между проводниками необходимо соблюдать промежуток не меньше 5 см, изгибы провода не должны касаться друг с другом.
- Также не рекомендуется допускать соприкосновение провода с арматурой, однако такое условие на практике не выполняется.
- Для более качественного прогрева провод следует укрыть слоем фольги.
- Подключение к трансформатору производится не греющим проводом, а через обычный провод. Обычно для таких целей применяют провод ПВ1, являющийся аналогом греющего проводника.

Провод ПВ1 и греющий ПНСВ должны быть соединены внутри бетона. При этом каких-либо специальных требований к качеству подключения не предъявляется. Соединение проводов должно производиться, как любое соединение электрических проводников, соответствующее правилам устройства и эксплуатации, и способное создать хорошую защиту от влаги.
Греющая проволока для прогревания бетона за приемлемую стоимость дает возможность решать проблемы с работами по заливке бетона в холодный период времени. При этом нет особых требований по эксплуатации и подключению.
При этом нет особых требований по эксплуатации и подключению.
Основным затруднением является расчет требуемой длины и подбор нужной схемы подключения. Но небольшие недочеты на этом этапе можно компенсировать, изменив рабочее напряжение, подаваемое в цепь.
Похожие темы:
Провод ПНСВ | Расшифровка, технические характеристики, описание
Расшифровка маркировки
- П — расшифровка позволяет определить вид кабельных изделий как провод;
- Н — назначение — для нагрева;
- С — материал изготовления жилы — сталь;
- В — поливинилхлоридная изоляция.
Элементы конструкции
- Однопроволочная круглая стальная жила.
- Изоляционный слой выполнен из полиэтилена или ПВХ пластиката
Технические и эксплуатационные параметры
Технические характеристики изделия соответствуют заявленным при эксплуатации в температурном диапазоне от –60 до +50 °С. Наибольшая температура в эксплуатационный период может равняться +80 °С.
Прокладку кабеля допускается проводить при температуре окружающей среды выше –15 °С. В ходе монтажа требуется следить, чтобы радиус его изгиба был равен 5 наружным диаметрам и более. Минимальное значение — 25 мм. Проложенные проводники должны находиться на расстоянии друг от друга минимум в 15 мм, не допускается их взаимное соприкасание или пересечение.
Изделия предназначены для работы как в повторно-кратковременном, так и в длительном режиме.
Подводить электропитание к секции нагрева необходимо «холодными» концами. Участки, где провод соединяется с «холодным» концом желательно вывести вне зоны обогрева. Соединение рекомендуется выполнять путем пайки с использованием медного проволочного бандажа, с помощью гильз или клеммных коробок. Кроме того, можно задействовать любой другой способ при условии, что будет обеспечена надежность соединения в период эксплуатации.
Чтобы тепловое поле было равномерным, смонтированную проводку следует покрывать фольгой из металла толщиной от 0,2 до 0,5 мм.
Возможно изготовление секций нагрева из 2 или 3 отрезков кабеля.
Электросопротивление изоляции изделий составляет минимум 1МОм на 1 км длины при 15-25 °С.
Срок эксплуатации по гарантии — 2 года.
Минимальный период службы — 16 лет.
Уточнить цены на провод ПНСВ, изучить технические характеристики, описание и сферы применения, рассчитать стоимость доставки в свой регион и заказать нужные маркоразмеры вы можете, выбрав необходимую марку в каталоге.
Провод ПНСВ, для прогрева бетона, греющий кабель ПНСВ, VSE-E, Киев Украина
Расшифровка провода ПНСВ
- П — провод
- Н — нагревательный
- С — стальная жила
- В — поливинилхлоридная оболочка
Нагревательный кабель для бетона: провод, нагревательный, стальная жила, ПВХ-оболочка.
Провод для прогрева бетона, описание марки ПНСВ
Греющий провод ПНСВ 1.2 – это особый вид кабельной продукции. Он используется исключительно в целях качественного и эффективного прогрева бетона, что делает его незаменимым при проведении строительных работ.
Особенности и преимущества прогрева бетона греющим кабелей, греющий провод ПНСВ.
Сегодня технология обогрева бетона нагревательными проводами (ПНСВ 1) стала широко распространена при возведении разнообразных монолитных сооружений. Укладывается продукция непосредственно перед заливанием бетонного раствора. После этого кабель покрывается специальным жидким составом. Как только бетонная смесь будет залита, процесс застывания пойдёт намного быстрее благодаря проводу ПНСВ. При этом структура никак не повредится и не изменится под воздействием температуры.
Тот же принцип применим и для любых других строительных смесей. Провод ПНСВ актуально использовать как в условиях бытового строительства, так и на строительных площадках
Греющий кабель ПНСВ характеристики
Чтобы обеспечить высокий уровень безопасности и избежать возможных пожаров, важно выбирать кабель с соответствующими параметрами сечения и уровня напряжения.
- диаметр: 1,2 мм;
- температура эксплуатации провода ПНСВ: -60… +50 °С;
- рабочий ток: от 14 до 16 Ампер;
- сопротивление ПНСВ: в среднем около 0,15 Ом/м;
- температура монтажа ПНСВ: -25… +50 °С.

Где купить провод ПНСВ?
Среди достоинств провода стоит отметить сильный изоляционный слой, предотвращающий риски возгорания, а также минимальную вероятность переломов проводников или перегибов. Для повышения уровня защиты можно использовать провод совместно с УЗО и трансформаторными подстанциями, которые позволяют контролировать температуру нагрева, осуществлять экономию электроэнергии и обеспечивать безопасность сотрудников.
Автор: МЕГА КАБЕЛЬ
Эту страничку дополнительно ищут в сети по запросу: пнсв 1, греющий провод пнсв, спираль для прогрева бетона, пнсв купить, характеристики, прогрев бетона кабелем пнсв.
ПНСВ — дешевый провод для прогрева бетона — цены, прайс-лист 2021
Расшифровка условного обозначения ПНСВ
- П – буква на первой позиции маркировки определяет тип изделия, как провод электрический;
- Н – буква второй позиции показывает назначение провода, как нагревательного, т.е. предназначенного для преобразования электрического тока в тепло;
- С – буква третьей позиции устанавливает материал жилы, в данном случае – это однопроволочная сталь;
- В – буква на 4-й позиции говорит о материале изоляции жилы, которая означает, что изоляция выполнена из ПВХ-пластиката.

Следующие позиции в маркировке провода занимают цифровые обозначения, определяющие количество жил и их диаметр в миллиметрах, например, ПНСВ 1х1,2 — это провод одножильный с диаметром жилы 1,2 мм. Изделие выпускается в соответствии с требованиями ТУ 16.К71-013-88.
Конструкция и принцип работы
ПНСВ имеет однопроволочную сплошную стальную оцинкованную жилу круглого сечения (1), которая покрыта снаружи слоем изоляции из поливинилхлоридного пластиката (2). Жила может иметь обычную или оцинкованную поверхность.
Принцип работы нагревательного провода основан на использовании физического эффекта Джоуля-Ленца, связанного с выделением тепла при протекании электрического тока через проводник.
Погонное сопротивление жилы, в зависимости от её диаметра, составляет 0,11-0,22 Ом/м. При этом удельная электрическая мощность провода равна 20 Вт/м. Изоляционный слой толщиной не менее 0,8 мм обладает диэлектрическим сопротивлением более 1 МОм/м и обеспечивает не только надёжную электрическую изоляцию от коротких замыканий, но и защиту от механических повреждений, влажности, хорошую теплоотдачу, устойчивость к действию солевых, кислотных и щелочных растворов.
Важно! Следует иметь в виду, что провод рассчитан на подключение в рабочей среде (бетон или другая смесь). Во избежание вывода из строя его нельзя подключать на воздухе.
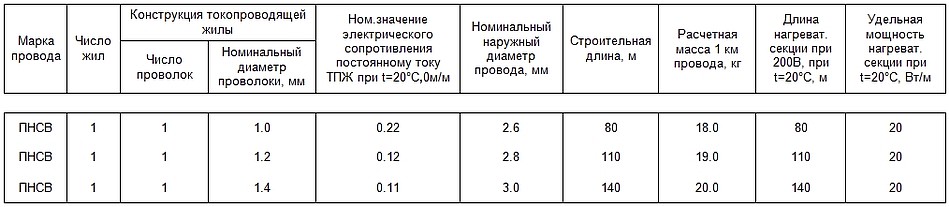
Для подключения следует применять «холодные концы» — отрезки провода типа АПВ-4. Провод можно эксплуатировать при температуре окружающей среды в пределах от -60°С до +80°С. Срок службы провода при соблюдении всех требований по эксплуатации составляет 16 лет.
Область применения греющего провода
Провод ПНСВ предназначен для нагрева бетонного раствора и обеспечения его быстрого затвердевания в зимних условиях. Основная сфера его применения – строительство. Для его питания используются специальные трансформаторы трёхфазного тока с напряжением 220/380В, при подключении к которым потребляемый рабочий ток равен 14-16 А.
Кроме того, провод эффективно применяется для обогрева производственных и бытовых помещений при использовании в качестве тёплых полов, для прогрева фундаментов, хранилищ продукции газовой и нефтеперерабатывающей промышленности. При этом для обеспечения равномерного распределения тепла, уложенные провода накрываются тонкой в 0,2-0,5 мм металлической фольгой. В данном случае питание ПНСВ производится от сети через выпрямитель переменного тока напряжением 60-80 В. При монтаже шаг укладки провода должен быть не менее 15 мм, а радиус изгиба не менее 25 мм.
При этом для обеспечения равномерного распределения тепла, уложенные провода накрываются тонкой в 0,2-0,5 мм металлической фольгой. В данном случае питание ПНСВ производится от сети через выпрямитель переменного тока напряжением 60-80 В. При монтаже шаг укладки провода должен быть не менее 15 мм, а радиус изгиба не менее 25 мм.
Провод ПНСВ: технические характеристики, расшифровка
Провод для прогрева бетона ПНСВ применяют для строительства различных объектов. Если сравнивать его с другими изделиями, данная модель не передаёт электрическую энергию, а отвечает за нагрев участка бетона, в который его нужно прокладывать.
Что это такое
Нагревательный кабель марки ПНСВ для прогрева бетона применяется в условиях затвердевания, происходящего при низких температурах. Он используется для напольных нагревателей с напряжением переменного тока до 380 В или постоянного тока до 1000 В.
Проводник ПНСВРасшифровка маркировки ПНСВ:
- «П» означает, что это провод, а не кабель;
- «Н» определяет основное назначение этого изделия и указывает, что этот проводник относится к нагревательным видам, и применять его для транспортировки электроэнергии не следует;
- «С» обозначает материал токоведущей жилы. Для этой марки таким материалом является сталь, которая обладает далеко не лучшим параметром теплопроводности, что и дает возможность использовать ее в качестве нагревательного элемента. Для производства токоведущих жил могут использоваться две категории стальной проволоки: оцинкованной и не имеющей покрытия. Оцинкованная жила защищена от появления ржавчины, но стоимость таких изделий выше;
- буква «В» означает, что материалом изоляции проводника является поливинилхлорид (ПВХ-оболочка).
 Для этой марки таким материалом является сталь, которая обладает далеко не лучшим параметром теплопроводности, что и дает возможность использовать ее в качестве нагревательного элемента. Для производства токоведущих жил могут использоваться две категории стальной проволоки: оцинкованной и не имеющей покрытия. Оцинкованная жила защищена от появления ржавчины, но стоимость таких изделий выше;
Для этой марки таким материалом является сталь, которая обладает далеко не лучшим параметром теплопроводности, что и дает возможность использовать ее в качестве нагревательного элемента. Для производства токоведущих жил могут использоваться две категории стальной проволоки: оцинкованной и не имеющей покрытия. Оцинкованная жила защищена от появления ржавчины, но стоимость таких изделий выше;Обратите внимание! В маркировке указано, из чего состоит провод: стальная жила, оцинкованная или не оцинкованная, покрыта изоляцией из ПВХ или специального полиэтилена.
Зачем нужен
Процесс, происходящий в бетоне в период его застывания, напрямую зависит от температурного режима. Зимой, когда температура значительно снижается, вода естественно замерзает. Это может привести к возникновению проблем с гидратацией бетона. Таким образом растров твердеет не полностью, так как на некоторых участках происходит его замерзание.
С приходом весны температурный режим постепенно повышается, вода тает, а цельность раствора нарушается. Это явление оказывает негативное влияние на целостность конструкции. В связи с этим значительно снижается её долговечность.
Обратите внимание! Чтобы предотвратить неблагоприятные последствия, необходимо обеспечить электропрогрев бетона проводами различного типа.
Важно отметить, что посторонний процесс в самом растворе не производится. Это свидетельствует о том, что структура однородная. Подобное явление оказывает благоприятное влияние на долговечность, а также прочность бетона.
Принцип работы
Стандартное время набора бетоном проектной прочности около 28 дней при температуре не ниже 15 ºС. В холодное время года процесс затвердевания бетонной смеси существенно замедляется.
Принцип действия кабеляЗамерзая и изменяя агрегатное состояние (превращаясь в лед), вода:
- не принимает участия в химических реакциях с цементом, поэтому процесс его гидратации замедляется, а сроки затвердевания бетона возрастают;
- существенно увеличивается в объеме (до 10 %), из-за чего возникает внутреннее давление, которое может деформировать или разрушить железобетонную конструкцию.

К сведению! Укладка бетонного раствора при низких и отрицательных температурах окружающей среды, если не принять необходимые меры, может привести к снижению качества и прочности бетона, что негативно сказывается на долговечности и безопасности железобетонных конструкций.
Технические характеристики
Характеристики греющего провода значительно отличаются от параметров обычных проводников, так как к нему предъявляются совсем другие требования. В этом виде проводов наиболее важными являются не свойства изоляции и токопроводящей жилы, а величина теплоотдачи и температурные свойства.
Технический показатель изделияЕсли рассматривать температурные параметры, то для провода ПНСВ обычно указывают допускаемую наибольшую температуру, величина которой 80 °С. При более высокой изоляция начинает разрушаться.
Обратите внимание! Эксплуатационная температура для этой марки проводника находится в интервале −60 °С — 50 °С.
Другой важной характеристикой является монтажная температура. Работа провода разрешается при морозах до −60 °С, но его укладка не должна производиться, если на улице ниже −15 °С.
Работа провода разрешается при морозах до −60 °С, но его укладка не должна производиться, если на улице ниже −15 °С.
Не менее важным параметром является удельная мощность выделения тепла. У проводников, предназначенных для передачи электричества, эта характеристика очень низкая. Для греющего провода производители указывают мощность выделения тепла около 20 Вт/м. Иногда изготовители заявляют этот параметр с величиной до 40 Вт/м, но здесь играет большую роль расчетная температура, а также наличие армирования поверхности, что значительно повышает этот параметр.
Следующим параметром греющего провода является его удельное сопротивление, которое прямо зависит от сечения токоведущей жилы. Например, расчет нагревательного провода марки ПНСВ с сечением 1,2 мм² обладает удельным сопротивлением 0,12 Ом/м, а провод 2 мм² — 0,044 Ом/м.
Если учесть, что этот вид проводника укладывается в бетонном растворе, то важным его свойством является устойчивость к проникновению воды, а также соляной и кислотной среды, что особенно важно для растворов, которые заливают при отрицательных температурах.
Важно! В такой бетон обычно добавляют разные присадки для достижения необходимой вязкости.
Преимущества и недостатки
Особенность сегментированного кабеля в том, что не нужно дополнительно включать устройство в систему. Подобный вариант прогрева бетона отличается максимальной безопасностью (сравнивая с электродами). Это характеризуется тем, что вероятность поражения электричеством исключается. Также выделяют другое преимущество — простоту монтажа и расчёта при эксплуатации нагревательной секции. Провод уже разбит на сегменты, остаётся лишь определить необходимую мощность.
Плюсы и минусы такого проводаОбратите внимание! Прогревать бетон зимой проводом ПНСВ гораздо дешевле. Поэтому сегментированный кабель, разбитый на сектора шинопроводов, используют лишь на некоторых объектах, где на первом месте маневренность возведения и точность работ.
Структура
Провода и кабели, особенно когда они зарыты в землю, требуют механической защиты от воздействия неблагоприятных условий окружающей среды, а также от неправильного обращения. На первый взгляд, все провода выглядят одинаково, но на самом деле существуют огромные различия. ПНСВ — провода для прогрева бетона, это их главная функция.
На первый взгляд, все провода выглядят одинаково, но на самом деле существуют огромные различия. ПНСВ — провода для прогрева бетона, это их главная функция.
Провода, используемые для ландшафтного и газонного орошения, обычно медные и изолированы различными материалами. Проводник может представлять собой одну жилу из твердого металла или несколько скрученных вместе жил. Многожильные проводники являются более гибкими и могут изгибаться взад и вперед во много раз больше, чем одножильный провод одинакового размера.
Элементы устройстваЭлектрическая изоляция должна выполнять двойную функцию: обеспечить электрическое сопротивление для защиты проводника от окружающей среды и механическую прочность, необходимую для поддержания его целостности во время монтажа и укладки траншеи.
Однако кабель для прогрева бетона ПНСВ имеет определённые минусы:
- сложно выполнять проектные работы, так как требуется точный расчёт длины изделия;
- нужно включать в систему понижающей подстанции;
- цена такой подстанций (ПТ) завышена;
- он должен регулярно выполнять свои функции, поэтому брать устройство в аренду нет смысла.

Обратите внимание! В ходе монтажа допустимо пользоваться сварочным аппаратом. Подобное устройство стоит применять для небольших объектов строительства. Из-за интенсивной работы устройство может выйти из строя.
Область применения
ПНСВ может использоваться для обогрева. В сельском хозяйстве и быту проводник применяется для обогрева почвы, водопроводных труб и желобов, для канализации, чтобы вода не замерзала в холодное время года. В помещениях провод нашел своё применение в качестве греющего элемента теплого электрического пола. Как уже было сказано, ПНСВ бывает двух видов: с оцинкованной и не оцинкованной жилой.
Область примененияВажно! Не оцинкованная жила подвержена коррозии.
Как правильно подключать
Для самостоятельного выполнения процедуры необходимо соблюдать определенные правила, так как для получения хорошего результата монтаж следует делать не на глаз, а путем точных расчетов.
ПодключениеОбратите внимание! Следует учесть объем бетона, площадь работ, температурный режим, скорость ветра в районе производства бетонных работ, необходимое время прогревания бетона, а также электрическую схему подключения.
Особенности подключения:
- в самом начале производится расчет необходимого количества провода, зависящего от рассмотренных выше параметров;
- провод ПНСВ следует подключать отдельными секциями. Подключение одним куском нецелесообразно, так как это не дает ожидаемого эффекта;
- вид подключения может быть трехфазным или двухфазным, а также по схемам треугольника или звезды;
- важными в ходе работ являются подбор длины провода и соблюдение нагрузки по току. На отдельной секции сила тока не должна превышать 15 Ам;
- соответствие подаваемого напряжения температуре провода. Чаще всего подключают питание в пределах 70-100 Вт, применяя для этого понижающий трансформатор. Наиболее популярной моделью трансформатора при проведении бетонных работ с использованием греющего провода стала марка КТПТО-80.
ПНСВ — провод для прогрева бетона, который за вполне приемлемую цену позволяет решать проблемы, связанные с бетонными работами в зимнее время. При этом он не предъявляет особых требований по подключению и эксплуатации.
При этом он не предъявляет особых требований по подключению и эксплуатации.
Провод пнсв или провод для прогрева бетона
В большинстве случаев воздействие положительных температур на бетонный раствор, находящийся в опалубке, организовывается при минусовых наружных температурах на стройплощадке. Но практика показывает, что прогревать бетон полезно и в других случаях – чтобы ускорить схватывание и затвердевание массы и получить максимально однородный состав. Один из таких методов, который называют активным прогреванием, использует провод для бетона, закладываемый непосредственно в рабочую массу. Укладка греющего кабеля в опалубку
Методика прогрева железобетона
Главное преимущество этой технологии перед другими методиками прогрева – абсолютное отсутствие потерь тепла, так как тепловая энергия остается в окружающей ее бетонной массе. Затраты же на нагревание кабеля и передачу тепла в тело бетонной конструкции – минимальные по сравнению с другими обогревающими технологиями.
Затраты же на нагревание кабеля и передачу тепла в тело бетонной конструкции – минимальные по сравнению с другими обогревающими технологиями.
Следующий, несомненно, большой плюс – простота реализации этого способа. Достаточно после расчетов правильно подобрать греющий элемент, схему подключения и укладки, выбрать подходящее напряжение, и конечного результата можно добиться, не прибегая к услугам строителей и электриков.
Сама технология состоит из нескольких этапов, первый из которых – сооружение опалубки или формы соответствующей конструкции, в которую будет укладывать армокаркас, греющий провод и будет заливаться бетон, который перед подачей напряжения в схему обязательно должен уплотняться глубинным вибратором. Заливка раствора в подготовленную форму
Некоторые особенности, которыми обладает ПНСВ или КДБС, использующиеся для прогрева бетонной массы, позволяют схеме более эффективно преобразовывать электроэнергию напряжения в тепловую, главное – правильно все рассчитать. Это тепло и греет бетон, ускоряя его схватывание и затвердевание.
Это тепло и греет бетон, ускоряя его схватывание и затвердевание.
Профессиональные строители знают и понимают разницу между греющим проводом и кабелем, а индивидуальным застройщикам будет полезно иметь об этом представление, что поможет использовать технологию правильно и более эффективно. Методика прогрева при помощи кабеля дороже по следующим причинам:
- Прогрев армированного бетона проводится при подключении схемы к пониженному напряжению через специальный понижающий трансформатор, чтобы не допустить при случайном повреждении изоляции утечки опасного напряжения в конструкцию через влажную металлическую арматуру.
- Греющий КДБС можно подключать к сети не менее 220в. Оптимально – 220в или 380в без понижения сетевого напряжения.
Подключить ПНСВ или КДБС проще в смысле организации рабочего процесса – соединения делаются через специальные муфты, а кабель при этом не нужно укорачивать или наращивать. Но греющий кабель дороже провода, поэтому в индивидуальном строительстве он используется на усмотрение хозяина. Еще один недостаток кабелей – их нельзя использовать повторно.
КДБС для прогрева бетона
Еще один недостаток кабелей – их нельзя использовать повторно.
КДБС для прогрева бетона
Отличия проводов:
- Внешне провод от кабеля можно отличить по следующим признакам: у провода обычно одна жила, у кабеля – две или больше.
- У провода номинальные температурные пределы при нагреве бетона – ± 55°С.
- Максимальная сила тока – 16 А.
- Сечение – 0,6-3 мм, что позволяет гнуть провод в любом направлении и использовать сложные схемы укладки. Можно использовать специальный калькулятор расчета сечения провода.
- Расход провода на нагревание 1 м3 бетонного раствора – 50-55 м.
Какие бывают греющие провода
ПНСВ 1.2 считается самой дешевой продукцией для прогревающих схем. Расшифровка характеристики ПНСВ: ПН – провод нагревательный, С – сталь, В – виниловая изоляция. Как уже говорилось, это самый дешевый вариант из существующих, поэтому в индивидуальном строительстве он более популярен, но для его эксплуатации нужен понижающий трансформатор, и в частном хозяйстве в этой роли может выступать обычный сварочный аппарат. ПНСВ 1.2
ПНСВ 1.2
Провод ПНСВ 2х1.2 представляет собой одну стальную жилу круглого сечения, с ПВХ или пластиковой изоляцией толщиной ≤ 0,8 мм. Рабочее напряжение для запитывания схемы с этим проводом – от 50 до 1000 вольт. Ток – переменный или постоянный. Возможность подключения к источнику постоянного тока позволяет подключать схему прогрева через понижающие трансформаторы с различным напряжением на выходе, но в пределах указанного выше значения. Электрическая развязка через трансформатор – это одна из мер безопасности при работе с высоким напряжением. Расчет температурного диапазона этой марки провода – от -60°С до +50°С, максимально допустимая – +80°С.
Почему эта марка так популярна у строителей и частных хозяйственников:
- Низкая возможность механического повреждения изоляции.
- При перепадах напряжения в сети, даже при подключении через понижающий трансформатор, провод не будет перегреваться так интенсивно, как аналоги.
- Из-за использования стальной жилы провод не деформируется в бетонной массе при ее схватывании и дальнейшем затвердевании.
- Сопротивление провода – 0,15 Ом/метр.
- Номинальная мощность – до 2,5 кВт/м3.
- Расчет расхода для 1 м3 бетонного раствора – до 60 погонных метров.
- Время схватывания бетона при использовании данного метода – до 72 часов.
Вопреки расхожему мнению, ПТПЖ 1х1.2, 2х1.2 (расшифровка: П – провод, Т – радиотрансляционный, П – изоляция из пластмассы, Ж – жила из оцинкованной стали) – это не кабель, а провод, который использовался в народно-хозяйственной промышленности еще до изобретения технологии подогрева бетона электричеством. Технические характеристики практически аналогичны параметрам ПНСВ, стальная жила бывает оцинкованной, сечение – 0,6-1,2 мм, изоляция ПЭВД (высокого давления). Единственное различие – в количестве жил: у ПТПЖ их две.
- ПТПЖ сохраняет рабочие характеристике при уличных температурах до -30°C.
- Радиус изгиба в схеме – не менее 10 диаметров жилы, чтобы не допустить появления микротрещин в изоляции.

Более экономная схема прогревания бетона с использованием ПТПЖ получится, если провод будет иметь сечение не более 0,6 мм. Также ПТПЖ используют в системе «теплый пол».
Также может быть использован АПВ (расшифровка: А – алюминиевая жила; П – провод; В – изоляция из ПВХ-пластиката (винила)). Шаг укладки провода в разных схемах
Особенности укладки греющего провода
Для каждой бетонной конструкции нужно разработать свою схему и подобрать соответствующие материалы, чтобы прогрев раствора проходил равномерно и сохранялась однородность структуры массы.
- Кабель подключается непосредственно к источнику напряжения, провод должен иметь так называемые «холодные» концы, удельное сопротивление (ρ) которых должно быть меньше ρ провода в схеме.
- Минимальный шаг размещения проводов вдоль длинной стороны формы – 15 мм. Их сближение может вызвать оплавление изоляции и короткое замыкание. Также нельзя накладывать их друг на друга.
- Хоть в спецификации к проводу и указаны допустимые диапазоны, на практике производить укладку при температуре на улице ниже -15°С не рекомендуется, так как увеличивается риск растрескивания изоляции, что может привести к КЗ.
- Эффект от обогрева можно увеличить, если поместить его в фольгу. Такая теплоизоляция повысит теплообмен и уменьшит сроки дозревания бетона до нормативных значений прочности.
Методика прогрева и укладки провода
Подготовительные работы, которые проводятся перед монтажом схемы:
- Сборка опалубки и армирующего каркаса. Все элементы этих конструкций должны быть освобождены от наледи.
- Провод укладывается на одном уровне с верхним и нижним рядами армирующего каркаса, без натяжения и сильного провисания, шаг укладки – 80-200 мм, конкретное расстояние зависит от погодных условий и уличной температуры. Пересечений и соприкосновений провода допускать нельзя, крепить кабель к арматуре следует пластиковыми хомутами, в крайнем случае – проволокой в изоляции или металлическими скрепками.
 Пересечений и соприкосновений провода допускать нельзя, крепить кабель к арматуре следует пластиковыми хомутами, в крайнем случае – проволокой в изоляции или металлическими скрепками.
Пересечений и соприкосновений провода допускать нельзя, крепить кабель к арматуре следует пластиковыми хомутами, в крайнем случае – проволокой в изоляции или металлическими скрепками.Калькулятор расчета
| Температура на улице, °С | Расстояние между проводом, см | Ø ПНСВ, мм | |
| для верхнего и нижнего ряда арматуры | для нижнего ряда арматуры | ||
| -5,0 | 20 | 10 | 1.1; 1.2; 1.4 |
| -10 | 16 | 8 | 1.1; 1.2; 1.4 |
| -15,0 | 12 | 8 | 1.1; 1.2; 1.4 |
| -20,0 | 10 | 8 | 1.1; 1.2; 1.4 |
- Понижающий трансформатор должен находиться от стройплощадки на расстоянии ≥ 25 м.
- Вокруг участка, на котором будет производиться бетонирование и обогрев раствора, устанавливается ограждение.
- ПНСВ подключается к секциям шин, к которым подключено питание от трансформатора.
- Шинопровод подключается к трансформатору, производится пробный холостой запуск электрической схемы на предмет проверки правильности сборки.

Схема должна учитывать реальное время прогревания бетонной смеси:
- Первоначальный период – это разогрев. Увеличение температуры в этом временном отрезке должно быть в пределах 10°С за 120 минут.
- Основной рабочий период – нагревание, при котором нельзя греть провод выше 80°С.
- Последний период работы схемы – остывание. В это время бетон должен остывать со скоростью ≤ 5°С в час.
Прогревание бетонной массы необходимо закончить после набора начальной прочности в пределах 50% от номинальной. При этом оптимальное время подогрева бетона будет варьироваться от 2-4 часов до трех суток – зависит от практических факторов: объема бетона, уличной температуры, схемы укладки и т.д.
 youtube.com/embed/i8C3kwj0eLY?feature=oembed&wmode=opaque» frameborder=»0″ allow=»accelerometer; autoplay; encrypted-media; gyroscope; picture-in-picture» allowfullscreen=»»/>
youtube.com/embed/i8C3kwj0eLY?feature=oembed&wmode=opaque» frameborder=»0″ allow=»accelerometer; autoplay; encrypted-media; gyroscope; picture-in-picture» allowfullscreen=»»/>EVM0008516.1 | Коттонген
Отношения
Эта мРНК является частью следующего гена . особенность (и):
Эта мРНК является членом следующей функции (ей) syntenic_region :
Следующие функции мРНК являются ортологами этой мРНК :
Следующие свойства полипептида происходят от этой мРНК:
Следующие функции CDS являются частью этой мРНК:
| Название функции | Уникальное имя | Виды | Тип |
|---|---|---|---|
| EVM0008516.1_D3d_CRI_v1-CDS-Chr09_D3d_CRI_v1-1627076778: 12968931..12969078 | EVM0008516.1_D3d_CRI_v1-CDS-Chr09_D3d_CRI_v1-1627076778: 12968931..12969078 | Gossypium davidsonii | |
| EVM0008516. 1_D3d_CRI_v1-CDS-Chr09_D3d_CRI_v1-1627076778 : 12969758..12970572 | EVM0008516.1_D3d_CRI_v1-CDS-Chr09_D3d_CRI_v1-1627076778: 12969758..12970572 | Gossypium davidsonii |
0
00 mdavidsonii
CDS 963 2 CDS 963 2 :
| Название функции | Уникальное имя | Виды | Тип |
|---|---|---|---|
| EVM0008516.1_D3d_CRI_v1-экзон-Chr09_D3d_CRI_v1-1627076778: 12968931..12969078 | EVM0008516.1_D3d_CRI_v1-экзон-Chr09_D3d_CRI_v1-1627076778: 12968931..12969078 | Gossypium davidsonii | |
| EVM0008516.1_D3d_CRI_v1-экзон-Chr09_D3d_CRI_v1-1627076778 : 12969758..12970572 | EVM0008516.1_D3d_CRI_v1-exon-Chr09_D3d_CRI_v1-1627076778: 12969758..12970572 | Gossypium davidsonii | экзон | Анализы Эта мРНК получена или имеет результаты следующих анализов Раскладки Следующие элементы выровнены Гомология ВЗРЫВ EVM0008516. 1 по сравнению с NCBI № 1 по сравнению с NCBI № Соответствие: gi | 10269 | ref | XP_016684316.1 | (ПРОГНОЗИРОВАННЫЙ: двухпоровый калиевый канал, подобный 3 [Gossypium hirsutum]) HSP 1 Оценка: 619,387 бит (1596), Expect = 0,000e + 0
Запрос: 1 MGTNLHRSKTAPAIAVLNDIIHPTIPKPQFGSPSIVRQAFVLVLLYLSLGVIIYWFNRSNFVGHETHPIVDVPYFCIVTICTIRYGDITPNSVTTKLFSILFVLVGFGFIDILL-GMISYVLDLQENYLLRTVQHGGQKKDSTRSYIIDVKKGRMRIRMKVGLALGVVVLCIGIGVGAMHFVESLGWLDSFYLSVMSVTTVGYGDRVFKTLPGRLFATIWLFVSTLAVARAFLYLAEARVGKRHRKMTKWVLGHDLTISEFLPADIDNNGFVSKLEFVIYKLKEMGKVSEKDIMLICETFDRLDVGNCGKITLADLVESHH 320
MGTNLHRSKTAPAIAVLNDIIHPTIPKPQFGSPSIVRQAFVLVLLYLSLGVIIYWFNRSNFVGHETHPIVDV YFCIVTICTI YGDITPNSVTTKLFSILFVLVGFGFID + LL GMISYVLDLQENYLLRTVQHGGQKKDSTRSYIIDVKKGRMRIRMK + GLALGVVVLCIGIGVGAMHFVESLGWLDSFYLSVMSVTTVGYGDRVFKTLPGRLFATIWLFVSTLAVARAFLYLAEARVGKRHR WVLGHDLTISEFL ADIDNNGFVSKLEFVIYKLKE + GKVSEKDIM ICETFDRLDVGNCGKITLADLVESHH
Sbjct: 1 MGTNLHRSKTAPAIAVLNDIIHPTIPKPQFGSPSIVRQAFVLVLLYLSLGVIIYWFNRSNFVGHETHPIVDVLYFCIVTICTIGYGDITPNSVTTKLFSILFVLVGFGFIDMLLSGMISYVLDLQENYLLRTVQHGGQKKDSTRSYIIDVKKGRMRIRMKMGLALGVVVLCIGIGVGAMHFVESLGWLDSFYLSVMSVTTVGYGDRVFKTLPGRLFATIWLFVSTLAVARAFLYLAEARVGKRHR ---- WVLGHDLTISEFLAADIDNNGFVSKLEFVIYKLKELGKVSEKDIMQICETFDRLDVGNCGKITLADLVESHH 317 ВЗРЫВ EVM0008516. 1 по сравнению с NCBI № 1 по сравнению с NCBI № Соответствие: gi | 1204924089 | ref | XP_021295064.1 | (двухпоровый калиевый канал 3-подобный [Herrania umbratica]) HSP 1 Оценка: 569,696 бит (1467), Expect = 0,000e + 0
Запрос: 3 TNLHRSKTAPAIAVLNDIIHPTIPKPQFGSPSIVRQAFVLVLLYLSLGVIIYWFNRSNFVGHETHPIVDVPYFCIVTICTIRYGDITPNSVTTKLFSILFVLVGFGFIDILL-GMISYVLDLQENYLLRTVQHGGQKKDSTRSYIIDVKKGRMRIRMKVGLALGVVVLCIGIGVGAMHFVESLGWLDSFYLSVMSVTTVGYGDRVFKTLPGRLFATIWLFVSTLAVARAFLYLAEARVGKRHRKMTKWVLGHDLTISEFLPADIDNNGFVSKLEFVIYKLKEMGKVSEKDIMLICETFDRLDVGNCGKITLADLVESHH 320
TNLHRSKTAPA + AVLND ++ HP + IPKPQFGS SIVRQAF + L ++ LYLSLGV + IYWFNR нелфинавир HETHP + В.Д. YFCIVT + ИТК YGDITPNSV TKLFSILFVLVGFGFIDILL GM + SYVLDLQENYLLRTV + G + K DS RSYIIDVKKGRMRIRMKVGLALGVVVLCIGIGVG MHFVESLGWLDSFYLSVMSVTTVGYGDR FKTLPGR + FA IWL VSTLAVARAFLYLAEARV KRHR + М KWVLGHD + T ++ EFL AD + DNNGFVSK EFVIYKLKEMGKVSEKDI + ICE FDRLD G + CGKITLADL + ESHH
Sbjct: 118 TNLHRSKTAPAMAVLNDVLHPSIPKPQFGSQSIVRQAFILLVLYLSLGVVIYWFNRRNFVAHETHPVVDALYFCIVTMCTIGYGDITPNSVATKLFSILFVLVGFGFIDILLTGMVSYVLDLQENYLLRTVKPEGEK-DSARSYIIDVKKGRMRIRMKVGLALGVVVLCIGIGVGVMHFVESLGWLDSFYLSVMSVTTVGYGDRAFKTLPGRIFAAIWLLVSTLAVARAFLYLAEARVDKRHRRMAKWVLGHDMTVAEFLAADMDNNGFVSKSEFVIYKLKEMGKVSEKDILQICEKFDRLDAGSCGKITLADLMESHH 435 ВЗРЫВ EVM0008516. 1 по сравнению с NCBI № 1 по сравнению с NCBI № Соответствие: gi | 1063511691 | ref | XP_017979162.1 | (ПРОГНОЗ: двухпоровый калиевый канал 3 [Theobroma cacao]> gi | 1063511693 | ref | XP_017979163.1 | ПРОГНОЗ: двухпоровый калиевый канал 3 [Theobroma cacao]) HSP 1 Оценка: 565,074 бит (1455), Ожидание = 0,000e + 0
Запрос: 3 TNLHRSKTAPAIAVLNDIIHPTIPKPQFGSPSIVRQAFVLVLLYLSLGVIIYWFNRSNFVGHETHPIVDVPYFCIVTICTIRYGDITPNSVTTKLFSILFVLVGFGFIDILL-GMISYVLDLQENYLLRTVQHGGQKKDSTRSYIIDVKKGRMRIRMKVGLALGVVVLCIGIGVGAMHFVESLGWLDSFYLSVMSVTTVGYGDRVFKTLPGRLFATIWLFVSTLAVARAFLYLAEARVGKRHRKMTKWVLGHDLTISEFLPADIDNNGFVSKLEFVIYKLKEMGKVSEKDIMLICETFDRLDVGNCGKITLADLVESHH 320
TNLHRSKTAPA + AVLND ++ HP + IPKPQFGS S + VRQAF + L ++ LYLSLGV + IYWFNR нелфинавир + ETHP + В.Д. YFCIVT + ИТК YGDITPNSV TKLFSILFVLVGFGFIDILL GM + SYVLDLQENYLLRTV + G + K DS RSYIIDVKKGRMRIRMKVGLALGVVVLCIGIGVG MHFVE + LGWLDSFYLSVMSVTTVGYGDR FKTLPGR + FA IWL VSTLAVARAFLYLAEARV KRHR М KWVLG D + Т ++ EFL ADIDNNGFVSK EFVIYKLKEMGKVSEKDI + ICE FDRLD GNCGKITLADL + ESHH
Sbjct: 119 TNLHRSKTAPAMAVLNDVLHPSIPKPQFGSQSVVRQAFILLVLYLSLGVVIYWFNRHNFVAYETHPVVDALYFCIVTMCTIGYGDITPNSVATKLFSILFVLVGFGFIDILLSGMVSYVLDLQENYLLRTVKPEGEK-DSARSYIIDVKKGRMRIRMKVGLALGVVVLCIGIGVGVMHFVETLGWLDSFYLSVMSVTTVGYGDRAFKTLPGRIFAAIWLLVSTLAVARAFLYLAEARVDKRHRTMAKWVLGQDMTVAEFLAADIDNNGFVSKSEFVIYKLKEMGKVSEKDILQICEKFDRLDAGNCGKITLADLMESHH 436 ВЗРЫВ EVM0008516. 1 по сравнению с NCBI № 1 по сравнению с NCBI № Соответствие: gi | 1269932939 | ref | XP_022715625.1 | (двухпоровый калиевый канал 3-подобный [Durio zibethinus]) HSP 1 Оценка: 564,303 бит (1453), Expect = 0,000e + 0
Запрос: 3 TNLHRSKTAPAIAVLNDIIHPTIPKPQFGSPSIVRQAFVLVLLYLSLGVIIYWFNRSNFVGHETHPIVDVPYFCIVTICTIRYGDITPNSVTTKLFSILFVLVGFGFIDILL-GMISYVLDLQENYLLRTVQHGGQKKDSTRSYIIDVKKGRMRIRMKVGLALGVVVLCIGIGVGAMHFVESLGWLDSFYLSVMSVTTVGYGDRVFKTLPGRLFATIWLFVSTLAVARAFLYLAEARVGKRHRKMTKWVLGHDLTISEFLPADIDNNGFVSKLEFVIYKLKEMGKVSEKDIMLICETFDRLDVGNCGKITLADLVESHH 320
TNLHRSKTAPA + AVLNDIIHP + IPKP FGS SIVRQAFVL ++ LYLSLGV + IYWFNR NFVGH ТНР + В.Д. YFCIVT + ИТК YGDITPNSV TKLFSILFVLVGFGFIDILL GM + SYVLDLQENYLLRTVQ G + K + DS RSYIIDVKKGRMRIRMKVGLALGVVVLCIGIG G MHFVESLGWLD + FYLSVMSVTTVGYGD FKTLPGR + FA IWL VSTLAVARAFLYLAEARV KRHR + М KWVLG D + Т + SEFL ADIDNNGFVSK Е.Ф. + IYKLKEMGKVSEKDIM IE FDRLD G CGKITLADL + ESHH
Sbjct: 118 TNLHRSKTAPAMAVLNDIIHPSIPKPPFGSQSIVRQAFVLLVLYLSLGVVIYWFNRHNFVGHVTHPVVDALYFCIVTMCTIGYGDITPNSVATKLFSILFVLVGFGFIDILLSGMVSYVLDLQENYLLRTVQDEGKKRDSARSYIIDVKKGRMRIRMKVGLALGVVVLCIGIGSGVMHFVESLGWLDAFYLSVMSVTTVGYGDTAFKTLPGRIFAAIWLLVSTLAVARAFLYLAEARVDKRHRRMAKWVLGQDMTVSEFLAADIDNNGFVSKSEFLIYKLKEMGKVSEKDIMQISEKFDRLDAGKCGKITLADLMESHH 436 ВЗРЫВ EVM0008516. 1 по сравнению с NCBI № 1 по сравнению с NCBI № Соответствие: gi | 508780532 | gb | EOY27788.1 | (Ca2 + активировал выпрямляющую наружу изоформу 1 канала 6 K + [Theobroma cacao]) HSP 1 Оценка: 562,762 бит (1449), Expect = 0,000e + 0
Запрос: 3 TNLHRSKTAPAIAVLNDIIHPTIPKPQFGSPSIVRQAFVLVLLYLSLGVIIYWFNRSNFVGHETHPIVDVPYFCIVTICTIRYGDITPNSVTTKLFSILFVLVGFGFIDILL-GMISYVLDLQENYLLRTVQHGGQKKDSTRSYIIDVKKGRMRIRMKVGLALGVVVLCIGIGVGAMHFVESLGWLDSFYLSVMSVTTVGYGDRVFKTLPGRLFATIWLFVSTLAVARAFLYLAEARVGKRHRKMTKWVLGHDLTISEFLPADIDNNGFVSKLEFVIYKLKEMGKVSEKDIMLICETFDRLDVGNCGKITLADLVESHH 320
TNLHRSKTAPA + AVLND ++ HP + IPKPQFGS S + VRQAF + L ++ LYLSLGV + IYWFNR нелфинавир + ETHP + В.Д. YFCIVT + ИТК YGDITPNSV TKLFSILFVLVGFGFIDILL GM + SYVLDLQENYLLRTV + G + K DS RSYIIDVKKGRMRIRMKVGLALGVVVLCIGIGVG MHFVESLGWLDSFYLSVMSVTTVGYGDR FKTLPGR + FA IWL VSTLAVARAFLYLAEARV KRHR М KWVLG D + Т ++ EFL ADIDNNGFVSK EFVIYKLKEMGKVSEKDI + ICE FDRLD GNCGKITLADL + ESHH
Sbjct: 215 TNLHRSKTAPAMAVLNDVLHPSIPKPQFGSQSVVRQAFILLVLYLSLGVVIYWFNRHNFVAYETHPVVDALYFCIVTMCTIGYGDITPNSVATKLFSILFVLVGFGFIDILLSGMVSYVLDLQENYLLRTVKPEGEK-DSARSYIIDVKKGRMRIRMKVGLALGVVVLCIGIGVGVMHFVESLGWLDSFYLSVMSVTTVGYGDRAFKTLPGRIFAAIWLLVSTLAVARAFLYLAEARVDKRHRTMAKWVLGQDMTVAEFLAADIDNNGFVSKSEFVIYKLKEMGKVSEKDILQICEKFDRLDAGNCGKITLADLMESHH 532 ВЗРЫВ EVM0008516. 1 по сравнению с NCBI № 1 по сравнению с NCBI № Соответствие: gi | 1028997318 | ref | XP_016753435.1 | (ПРОГНОЗИРОВАННЫЙ: изоформа X1, подобная трехпорному калиевому каналу [Gossypium hirsutum]) HSP 1 Оценка: 558,14 бит (1437), Ожидаемое значение = 0,000e + 0
Запрос: 3 TNLHRSKTAPAIAVLNDIIHPTIPKPQFGSPSIVRQAFVLVLLYLSLGVIIYWFNRSNFVGHETHPIVDVPYFCIVTICTIRYGDITPNSVTTKLFSILFVLVGFGFIDILL-GMISYVLDLQENYLLRTVQHGGQKKDSTRSYIIDVKKGRMRIRMKVGLALGVVVLCIGIGVGAMHFVESLGWLDSFYLSVMSVTTVGYGDRVFKTLPGRLFATIWLFVSTLAVARAFLYLAEARVGKRHRKMTKWVLGHDLTISEFLPADIDNNGFVSKLEFVIYKLKEMGKVSEKDIMLICETFDRLDVGNCGKITLADLVESHH 320
+ NLHRSKTAPA + AVLND + I + IPKPQ Г.С. SIVRQAFVL ++ LYL + LGV + I + WFNR NF + G + ETHP + В.Д. YFCIVT + ИТК YGDI ПНСВ TKLFSILFVLVGFGFIDILL GM + SYVLDLQE + YLLRTV + Н GQKKDS RSYIIDVKKGRMRIRMKVGLALGVVVLCIGIGVG MHFVE LGWLD + FYLSVMSVTTVGYGDR FKTLPGR + FA IWL VSTLAVARAFLYLAEARV KRHR M KWVLG D + T + SEFL ADIDNNGFVSK EFVIYKLKEMGKVSEKDIM IE FDRLD GNCGKITLADL + E HH
Sbjct: 123 SNLHRSKTAPAMAVLNDVIRSSIPKPQLGSQSIVRQAFVLLVLYLTLGVVIFWFNRHNFLGNETHPVVDALYFCIVTMCTIGYGDIIPNSVATKLFSILFVLVGFGFIDILLSGMVSYVLDLQESYLLRTVEHEGQKKDSARSYIIDVKKGRMRIRMKVGLALGVVVLCIGIGVGVMHFVERLGWLDAFYLSVMSVTTVGYGDRAFKTLPGRIFAAIWLLVSTLAVARAFLYLAEARVDKRHRTMAKWVLGQDMTVSEFLAADIDNNGFVSKSEFVIYKLKEMGKVSEKDIMQISEKFDRLDAGNCGKITLADLMEHHH 441 ВЗРЫВ EVM0008516. 1 по сравнению с NCBI № 1 по сравнению с NCBI № Соответствие: gi | 1050646378 | ref | XP_017646501.1 | (ПРОГНОЗИРОВАННЫЙ: изоформа X1, подобная трехпорному калиевому каналу [Gossypium arboreum]) HSP 1 Оценка: 557,755 бит (1436), Ожидаемое значение = 0,000e + 0
Запрос: 3 TNLHRSKTAPAIAVLNDIIHPTIPKPQFGSPSIVRQAFVLVLLYLSLGVIIYWFNRSNFVGHETHPIVDVPYFCIVTICTIRYGDITPNSVTTKLFSILFVLVGFGFIDILL-GMISYVLDLQENYLLRTVQHGGQKKDSTRSYIIDVKKGRMRIRMKVGLALGVVVLCIGIGVGAMHFVESLGWLDSFYLSVMSVTTVGYGDRVFKTLPGRLFATIWLFVSTLAVARAFLYLAEARVGKRHRKMTKWVLGHDLTISEFLPADIDNNGFVSKLEFVIYKLKEMGKVSEKDIMLICETFDRLDVGNCGKITLADLVESHH 320
+ NLHRSKTAPA + AVLND + I + IPKPQ Г.С. SIVRQAFVL ++ LYL + LGV + I + WFNR NF + G + ETHP + В.Д. YFCIVT + ИТК YGDI ПНСВ TKLFSILFVLVGFGFIDILL GM + SYVLDLQE + YLLRTV + Н GQKKDS RSYIIDVKKGRMRIRMKVGLALGVVVLCIGIGVG MHFVE LGWLD + FYLSVMSVTTVGYGDR FKTLPGR + FA IWL VSTLAVARAFLYLAEARV KRHR M KWVLG D + T + SEFL ADIDNNGFVSK EFVIYKLKEMGKVSEKDIM IE FDRLD GNCGKITLADL + E HH
Sbjct: 123 SNLHRSKTAPAMAVLNDVIRSSIPKPQLGSQSIVRQAFVLLVLYLTLGVVIFWFNRHNFLGNETHPVVDALYFCIVTMCTIGYGDIIPNSVATKLFSILFVLVGFGFIDILLSGMVSYVLDLQESYLLRTVEHEGQKKDSARSYIIDVKKGRMRIRMKVGLALGVVVLCIGIGVGVMHFVERLGWLDAFYLSVMSVTTVGYGDRAFKTLPGRIFAAIWLLVSTLAVARAFLYLAEARVDKRHRTMAKWVLGQDMTVSEFLAADIDNNGFVSKSEFVIYKLKEMGKVSEKDIMQISEKFDRLDAGNCGKITLADLMEHHH 441 ВЗРЫВ EVM0008516.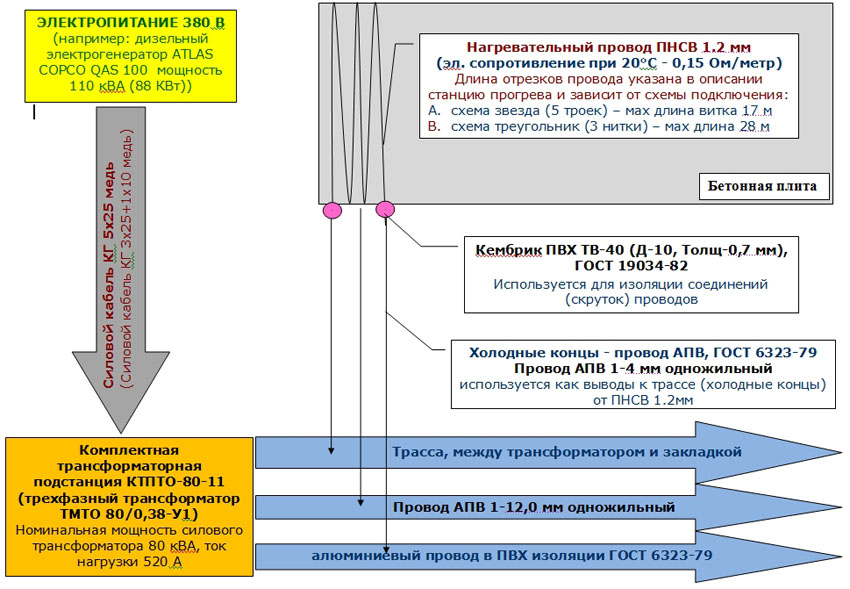 1 по сравнению с NCBI № 1 по сравнению с NCBI № Соответствие: gi | 1269948926 | ref | XP_022724189.1 | (двухпоровый калиевый канал 3-подобный [Durio zibethinus]) HSP 1 Оценка: 556,599 бит (1433), Expect = 0,000e + 0
Запрос: 3 TNLHRSKTAPAIAVLNDIIHPTIPKPQFGSPSIVRQAFVLVLLYLSLGVIIYWFNRSNFVGHETHPIVDVPYFCIVTICTIRYGDITPNSVTTKLFSILFVLVGFGFIDILL-GMISYVLDLQENYLLRTVQHGGQKKDSTRSYIIDVKKGRMRIRMKVGLALGVVVLCIGIGVGAMHFVESLGWLDSFYLSVMSVTTVGYGDRVFKTLPGRLFATIWLFVSTLAVARAFLYLAEARVGKRHRKMTKWVLGHDLTISEFLPADIDNNGFVSKLEFVIYKLKEMGKVSEKDIMLICETFDRLDVGNCGKITLADLVESHH 320
TNLHR KTAPA + AVLND МПЗ + IPKPQ Г.С. SIVRQAFVL ++ LYLSLGV + IYWFNR FVGHETHP + В.Д. МДХ + ВТ + ИТК YGDITPNSV TKLFSILFVLVGFGFIDILL GM + SYVLDLQENYLLRTVQ QKKDS RSYIIDVKKGRMRIRMKVGLALGVVVLCIGIGVG MHFVE + LGWLD + FYLSVMSVTTVGYGD F TLPGR + FA IWL VSTLAVARAFLYLAEARV KRHR + М KWVLG D + TISEFL ADIDNNGFVSK EFVIYKLKEMGKVSEKDIM И.Е. FDRLD GNCGKITL + L + ESHH
Sbjct: 182 TNLHRCKTAPAMAVLNDFIHPSIPKPQLGSQSIVRQAFVLLVLYLSLGVVIYWFNRHKFVGHETHPVVDGLYFCVVTMCTIGYGDITPNSVATKLFSILFVLVGFGFIDILLSGMVSYVLDLQENYLLRTVQQEVQKKDSARSYIIDVKKGRMRIRMKVGLALGVVVLCIGIGVGVMHFVENLGWLDAFYLSVMSVTTVGYGDAAFMTLPGRVFAAIWLLVSTLAVARAFLYLAEARVDKRHRRMAKWVLGQDMTISEFLAADIDNNGFVSKSEFVIYKLKEMGKVSEKDIMQISEKFDRLDAGNCGKITLVELMESHH 500 ВЗРЫВ EVM0008516. 1 по сравнению с NCBI № 1 по сравнению с NCBI № Соответствие: gi | 10284 | ref | XP_016683488.1 | (ПРОГНОЗИРОВАННЫЙ: изоформа X1, подобная трехпорному калиевому каналу [Gossypium hirsutum]) HSP 1 Оценка: 556,214 бит (1432), Ожидаемое значение = 0,000e + 0
Запрос: 3 TNLHRSKTAPAIAVLNDIIHPTIPKPQFGSPSIVRQAFVLVLLYLSLGVIIYWFNRSNFVGHETHPIVDVPYFCIVTICTIRYGDITPNSVTTKLFSILFVLVGFGFIDILL-GMISYVLDLQENYLLRTVQHGGQKKDSTRSYIIDVKKGRMRIRMKVGLALGVVVLCIGIGVGAMHFVESLGWLDSFYLSVMSVTTVGYGDRVFKTLPGRLFATIWLFVSTLAVARAFLYLAEARVGKRHRKMTKWVLGHDLTISEFLPADIDNNGFVSKLEFVIYKLKEMGKVSEKDIMLICETFDRLDVGNCGKITLADLVESHH 320
+ NLHRSKTAPA + AVLND + I + IPKPQ Г.С. SIVRQAFVL + LYL + LGV + I + WFNR NF + G + ETHP + В.Д. YFCIVT + ИТК YGDI ПНСВ TKLFSILFVLVGFGFIDILL GM + SYVLDLQE + YLLRTV + Н GQKKDS RSYIIDVKKGRMRIRMKVGLALGVVVLCIGIGVG MHFVE LGWLD + FYLSVMSVTTVGYGDR FKTLPGR + FA IWL VSTLAVARAFLYLAEARV KRHR М KWVLG D + T + SEFL ADIDNNGFVSK EFVIYKLKEMGKVSEKDIM IE FDRLD GNCGKITLADL + E HH
Sbjct: 123 SNLHRSKTAPAMAVLNDVIRSSIPKPQLGSQSIVRQAFVLLFLYLTLGVVIFWFNRHNFLGNETHPVVDALYFCIVTMCTIGYGDIIPNSVATKLFSILFVLVGFGFIDILLSGMVSYVLDLQESYLLRTVEHEGQKKDSARSYIIDVKKGRMRIRMKVGLALGVVVLCIGIGVGVMHFVERLGWLDAFYLSVMSVTTVGYGDRAFKTLPGRIFAAIWLLVSTLAVARAFLYLAEARVDKRHRTMAKWVLGQDMTVSEFLAADIDNNGFVSKSEFVIYKLKEMGKVSEKDIMQISEKFDRLDAGNCGKITLADLMEHHH 441 ВЗРЫВ EVM0008516.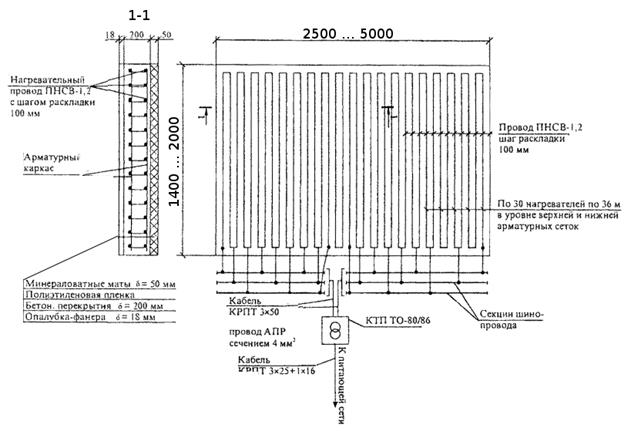 1 по сравнению с NCBI № 1 по сравнению с NCBI № Соответствие: gi | 823235220 | ref | XP_012450248.1 | (ПРОГНОЗИРОВАННЫЙ: изоформа X1, подобная двухпоровому калиевому каналу 3 [Gossypium raimondii]> gi | 763799957 | gb | KJB66912.1 | гипотетический белок B456_010G164600 [Gossypium raimondii]) HSP 146 Оценка: 55632 балла = 0,000e + 0
Запрос: 3 TNLHRSKTAPAIAVLNDIIHPTIPKPQFGSPSIVRQAFVLVLLYLSLGVIIYWFNRSNFVGHETHPIVDVPYFCIVTICTIRYGDITPNSVTTKLFSILFVLVGFGFIDILL-GMISYVLDLQENYLLRTVQHGGQKKDSTRSYIIDVKKGRMRIRMKVGLALGVVVLCIGIGVGAMHFVESLGWLDSFYLSVMSVTTVGYGDRVFKTLPGRLFATIWLFVSTLAVARAFLYLAEARVGKRHRKMTKWVLGHDLTISEFLPADIDNNGFVSKLEFVIYKLKEMGKVSEKDIMLICETFDRLDVGNCGKITLADLVESHH 320
+ NLHRSKTAPA + AVLND + I + IPKPQ Г.С. SIVRQAFVL ++ LYL + LGV + I + WFNR NF + G + ETHP + В.Д. YFCIVT + ИТК YGDI ПНСВ TKLFSILFVLVGFGFIDILL GM + SYVLDLQE + YLLRTV + Н GQKKDS RSYIIDVKKGRMRIRMKVGLALGVVVLCIGIGVG MHFVE LGWLD + FYLSVMSVTTVGYGDR FKTLPGR + FA IWL VSTLAVARAFLYLAEARV KRHR M KWVLG D + T + SEFL ADIDNNGFVSK EFVIYKLKEMGKVSEKDIM IE FDRLD GNCGKITLADL + E HH
Sbjct: 122 SNLHRSKTAPAMAVLNDVIRSSIPKPQLGSQSIVRQAFVLLVLYLTLGVVIFWFNRHNFLGNETHPVVDALYFCIVTMCTIGYGDIIPNSVATKLFSILFVLVGFGFIDILLSGMVSYVLDLQESYLLRTVEHEGQKKDSARSYIIDVKKGRMRIRMKVGLALGVVVLCIGIGVGVMHFVERLGWLDAFYLSVMSVTTVGYGDRAFKTLPGRIFAAIWLLVSTLAVARAFLYLAEARVDKRHRTMAKWVLGQDMTVSEFLAADIDNNGFVSKSEFVIYKLKEMGKVSEKDIMQISEKFDRLDAGNCGKITLADLMEHHH 440 ВЗРЫВ EVM0008516.1 в сравнении с ExPASy Swiss-Prot Соответствие: KCO6_ARATH (Двухпористый калиевый канал 3 OS = Arabidopsis thaliana OX = 3702 GN = TPK3 PE = 2 SV = 1) HSP 1 Оценка: 444,121 бит (1141), Expect = 4,951e-155
Запрос: 1 MGTNLHRSKTAPAIAVLNDIIHPTIPK-PQFGSPSIVRQAFVLVLLYLSLGVIIYWFNRSNFVGHETHPIVDVPYFCIVTICTIRYGDITPNSVTTKLFSILFVLVGFGFIDILL-GMISYVLDLQENYLLRTVQHGGQKKDSTRSYIIDVKKGRMRIRMKVGLALGVVVLCIGIGVGAMHFVESLGWLDSFYLSVMSVTTVGYGDRVFKTLPGRLFATIWLFVSTLAVARAFLYLAEARVGKRHRKMTKWVLGHDLTISEFLPADIDNNGFVSKLEFVIYKLKEMGKVSEKDIMLICETFDRLDVGNCGKITLADLVE 317
G LHRSKTAPA + AV + ND + HP KPS S + VRQAF L +++ YLSLGV + IYW NR ++ V ++ THP + VD YFCIVT + CTI YGDITPNSV TKLFSI + FVLVGFGFIDILL GM + SYVLDLQE + Y + L + V + KVGRLIDGI + V + KVGRI GVG MHF + E + GWLDSFYLSVMSVTTVGYGDR FKTLPGRLFA IWL VSTLAVARAFLYLAEARV KR + R + K VL +++ S + F ADIDNNG VSK E + VIYKLKEM K +++ KDI + I + FD + LD + E GKITL DL
Sbjct: 114 QGHALHRSKTAPAMAVINDLHHPIRQKDPTETSRSVVRQAFALLVVYLSLGVLIYWLNRDHYVVNQTHPVVDGLYFCIVTMCTIGYGDITPNSVVTKLFSIMFVLVGFGFIDILLSGMVSYVLDLQESYMLDSAKRRDEP-EKRRSYIIDVKKGRMRIRLKVALALGVVVLCIAVGVGIMHFIEEIGWLDSFYLSVMSVTTVGYGDRAFKTLPGRLFAAIWLLVSTLAVARAFLYLAEARVDKRNRERAKKVLCETMSVSQFFAADIDNNGCVSKAEYVIYKLKEMEKITDKDILPISKQFDKLDRCSNGKITLLDLLE 431 ВЗРЫВ EVM0008516.1 в сравнении с ExPASy Swiss-Prot Соответствие: KCO2_ARATH (Двухпористый калиевый канал 2 OS = Arabidopsis thaliana OX = 3702 GN = TPK2 PE = 2 SV = 1) HSP 1 Оценка: 429,098 бит (1102), Expect = 5,089e-149
Запрос: 4 NLHRSKTAPAIAVLNDIIHPTIPK - PQFGSPSIVRQAFVLVLLYLSLGVIIYWFNRSNFVGHETHPIVDVPYFCIVTICTIRYGDITPNSVTTKLFSILFVLVGFGFIDILL-GMISYVLDLQENYLLRTVQHGG --- QKKDSTRSYIIDVKKGRMRIRMKVGLALGVVVLCIGIGVGAMHFVESLGWLDSFYLSVMSVTTVGYGDRVFKTLPGRLFATIWLFVSTLAVARAFLYLAEARVGKRHRKMTKWVLGHDLTISEFLPADIDNNGFVSKLEFVIYKLKEMGKVSEKDIMLICETFDRLDVGNCGKITLADLVES 318
N HRSKTAPA + A + N + I HP PK QS + IV QA L +++ YLSLGV + IYW NR ++ + THP + VD YFCIVT + CTI YGDITP + SV TKLFSI FVLVGFGF + DILL GM ++ YVLDLQENY + LT ++ + DGRMRIRIDVKK KVGLALGVVVLC + G GV MHFVE + GWLDSFY SVMSVTTVGYGDR F TL GRL A + WL VSTLAVARA L + LAE + RV KR + R + K VLG ++ IS + FL ADID NG VSK EFVIYKLK + M K ++ EKDI I + LD + N ES
Sbjct: 112 NFHRSKTAPAMAAINNISHPNDPKTDQQSDSKTIVNQAVALLVVYLSLGVLIYWLNRDSYNVKQTHPVVDALYFCIVTMCTIGYGDITPDSVVTKLFSIFFVLVGFGFMDILLSGMVTYVLDLQENYMLETARNESLNLNDRDKVRSYIIDVKKGRMRIRLKVGLALGVVVLCLGFGVLIMHFVEKIGWLDSFYFSVMSVTTVGYGDRAFNTLAGRLLAAMWLLVSTLAVARAILFLAESRVDKRNRERAKKVLGESMSISQFLDADIDCNGCVSKAEFVIYKLKKMDKITEKDINPIGFQFDKLDRTNSGRITLLDLLES 432 ВЗРЫВ EVM0008516.1 в сравнении с ExPASy Swiss-Prot Соответствие: KCO3_ORYSJ (Двухпористый калиевый канал c OS = Oryza sativa subsp. Japonica OX = 39947 GN = TPKC PE = 3 SV = 1) HSP 1 Оценка: 396,741 бит (1018), Expect = 4,894e-136
Запрос: 3 TNLHRSKTAPAIAVLNDIIHPTIPKP -------- QFGSPSIVRQAFVLVLLYLSLGVIIYWFNRSNFVGHE - THPIVDVPYFCIVTICTIRYGDITPNSVTTKLFSILFVLVGFGFIDILL-GMISYVLDLQENYLLRTVQHGGQKKDSTRSYIIDVKKGRMRIRMKVGLALGVVVLCIGIGVGAMHFVESLGWLDSFYLSVMSVTTVGYGDRVFKTLPGRLFATIWLFVSTLAVARAFLYLAEARVGKRHRKMTKWVLGHDLTISEFLPADIDNNGFVSKLEFVIYKLKEMGKVSEKDIMLICETFDRLDVGNCGKITLADLVESHH 320
TNLHRS + TAPA + A LN P + IV AF + + L YL ++ GV Y NF THP + D YFCIVT + CTI YGDITP + KLFSI FVL + GFGF + DILL GM + SYVLDLQE + L + +++ + + YI D + KKGRMR + RMKV LAL VV + C + G + G + VE + LGWLD + YL + VMSVTTVGYGD F + TL GRLFA + WL VSTLAVARAFLYLAE R + KRHR M WVL D + T + SEFL ADIDNNG + V + K EFV + YKLKEMGK + SEKDIM + IC + F R + D + DLCG
Sbjct: 114 TNLHRSRTAPAMAPLNAAAIAAAAASGDSRNPPPPPRRPAIVLHAFLFLLAYLAMGVTFYAALPGNFTSSAGPTHPVADALYFCIVTLCTIGYGDITPATPAAKLFSISFVLIGFGFVDILLSGMVSYVLDLQEHLLITALKNPRSVRKHRHNYIFDLKKGRMRVRMKVALALTVVAICVGVGAAVLKRVENLGWLDAVYLAVMSVTTVGYGDHAFRTLAGRLFASAWLLVSTLAVARAFLYLAEMRIDKRHRAMANWVLSRDMTVSEFLAADIDNNGYVTKSEFVVYKLKEMGKISEKDIMMICDQFQRMDSGNCGKITLSDLLESHQ 442 ВЗРЫВ EVM0008516.1 в сравнении с ExPASy Swiss-Prot Соответствие: KCO5_ARATH (Двухпоровый калиевый канал 5 OS = Arabidopsis thaliana OX = 3702 GN = TPK5 PE = 1 SV = 1) HSP 1 Оценка: 366,696 бит (940), Expect = 5,960e-125
Запрос: 5 LHRSKTAPAIAVLNDI - IHPTIPKPQFGSPSIVRQAFVLVLLYLSLGVIIYWFNRSNFVGHETHPIVDVPYFCIVTICTIRYGDITPNSVTTKLFSILFVLVGFGFIDILL-GMISYVLDLQENYLLRTVQ ------ HGGQKKDSTRSYIIDVKKGRMRIRMKVGLALGVVVLCIGIGVGAMHFVESLGWLDSFYLSVMSVTTVGYGDRVFKTLPGRLFATIWLFVSTLAVARAFLYLAEARVGKRHRKMTKWVLGHDLTISEFLPADIDNNGFVSKLEFVIYKLKEMGKVSEKDIMLICETFDRLDVGNCGKITLADLV 316
LHRS + TAPA + ++ D + P KP S SI + RQA L +++ YL + LGV IY FNR ++ G ETHP + VD YFCIVT + CTI YGDI P + TK + F +++ FVL GFGF + DILL G +++ YVLDLQE + + L + QH + S + YIID + KGRMRIRMKV LAL VVVLCIG + G + HFVE LG ++ DS YLSVMSVTTVGYGDR FKTL GRLFA + WL VSTLAVARAFLYLAEAR + + RHRK KL ++ T + + L AD + GF + SK E + +++ KLKDI + KLKDI + KLKDI + KLKDI
Sbjct: 84 LHRSRTAPAMVIIKDLRTKPPETKKPSPVSKSIIRQAIFLLIVYLTLGVSIYSFNRDHYSGIETHPVVDALYFCIVTMCTIGYGDIAPLTPWTKIFAVVFVLFGFGFLDILLSGVVNYVLDLQESMILTGIQTRQHHQHHHHHRFSAKDYIIDFEKGRMRIRMKVCLALCVVVLCIGVGALVLHFVEELGFVDSVYLSVMSVTTVGYGDRAFKTLQGRLFAAVWLLVSTLAVARAFLYLAEARIDRRHRKAVKLALNREITVDDLLKADTYQHGFISKSEYIVLKLKEMGKITQKDIDQVVIQFEKLDPNQIGKITLPDLL 404 ВЗРЫВ EVM0008516.1 в сравнении с ExPASy Swiss-Prot Соответствие: KCO4_ARATH (Двухпоровый калиевый канал 4 OS = Arabidopsis thaliana OX = 3702 GN = TPK4 PE = 2 SV = 2) HSP 1 Оценка: 197,978 бит (502), Expect = 1,428e-60
Запрос: 22 HPTIPKPQFGSPSIVRQ -------- AFVLVLLYLSLGVIIYWFNRSNFVGHETHPIVDVPYFCIVTICTIRYGDITPNSVTTKLFSILFVLVGFGFIDILLG-MISYVLDLQENYLLRTVQHGGQKKDSTRSYIIDVKKGRMRIRMKVGLALGVVVLCIGIGVGAMHFVESLGWLDSFYLSVMSVTTVGYGDRVFKTLPGRLFATIWLFVSTLAVARAFLYLAEARVGKRHRKMTKWVLGHDLTISEFLPADIDNNGFVSKLEFVIYKLKEMGKVSEKDIMLICETFDRLD 303
HP P + + V + A + L + L + YL + GV YFRFG ET + VD YF IVT T + YGDI P ++ TTK + + I + VG F + D LL ++ S + VL LQEN + L + + + R + I + G ++ R ++ K + LA V LC + GG + HEL WLDS YLSV + SVTTVGYGD + FKT + GR FA WL + ST + A + A FLYLAE R + + T + + P S + EF +++ KL + E G ++ SE DI I F + L +
Sbjct: 13 HPNESSPEETQVTTVSKSKWTILVLAMILLLVYLTFGVCTYSFFRDQFSGTETNLFVDAFYFSIVTFSTVGYGDIVPSTSTTKILTIVLVSTGVVFLDYLLNRVVSHVLSLQENAILDRINK - TRNRAIRDHI - AEDGKIRLKWKLCLAFCAVGLCVGSGALFLHVFERLDWLDSVYLSVISVTTVGYGDKTFKTVEGRGFAVFWLLLSTIAMATLFLYLAEMRIDRT ------------- TVMKLPP --------- SESEFIVFKLRESGRISEDDIKQIVREFENLE 277 ВЗРЫВ EVM0008516.1 в сравнении с ExPASy Swiss-Prot Соответствие: KCO1_ARATH (Двухпоровый калиевый канал 1 OS = Arabidopsis thaliana OX = 3702 GN = TPK1 PE = 1 SV = 2) HSP 1 Оценка: 187,578 бит (475), Expect = 1,314e-55
Запрос: 22 HPT-IPKPQFGSPSIVRQAFVLVLLYLSLGVIIYWFNRSNFVGHETHPIVDVPYFCIVTICTIRYGDITPNSVTTKLFSILFVLVGFGFIDILLGMIS-YVLDLQENYLLRTVQHGGQKKDSTRSYIIDVKKG - RMRIRMKVGLALGVVVLCIGIGVGAMHFVESLGWLDSFYLSVMSVTTVGYGDRVFKTLPGRLFATIWLFVSTLAVARAFLYLAEARVGKRHRKMTKWVLGHDLTISEFLPADIDNNGFVSKLEFVIYKLKEMGKVSEKDIMLICETFDRLDVGNCGKITLADLV 316
HP + IP + P ++ R + + + LYL ++ G + ++ R GH + T + VD YFCIVT + T + YGD + PNS ++ L + FV G + LL + Y +++ QE L + RHQT D + K ++ RK V + V + + G + VE + + + FY + VTT + GYGD + F + GRLFA W + S ++ + A + FLY + AE + R + KWVL + T ++ AD + D + GV EF ++ YKLKEMGK + EKDI I + F ++ LD G + T + D + V
Sbjct: 62 HPSKIPMFSDLNPNL-RRVIMFLALYLTIGTLCFYLVRDQISGHKTSGVVDALYFCIVTMTTVGYGDLVPNSSASRLLACAFVFSGMVLVGHLLSRAADYLVEKQEALLVRAF-HLRQSFGPT ----- DILKELHTNKLRYKCYATCLVLVVLFIVGTIFLVMVEKMPVISAFYCVCSTVTTLGYGDKSFNSEAGRLFAVFWILTSSICLAQFFLYVAELNTENKQRALVKWVLTRRITNNDLEAADLDEDGVVGAAEFIVYKLKEMGKIDEKDISGIMDEFEQLDYDESGTLTTSDIV 353 ВЗРЫВ EVM0008516.1 в сравнении с ExPASy Swiss-Prot Соответствие: KCO3_ARATH (Калийный внутренний выпрямитель (Kir) -подобный канал 3 OS = Arabidopsis thaliana OX = 3702 GN = KCO3 PE = 1 SV = 1) HSP 1 Оценка: 171,4 бит (433), Expect = 1,354e- 50
Запрос: 160 VGLALGVVVLCIGIGVGAMHFVESLGWLDSFYLSVMSVTTVGYGDRVFKTLPGRLFATIWLFVSTLAVARAFLYLAEARVGKRHRKMTKWVLGHDLTISEFLPADIDNNGVIYFVSKLE
VL VV CG + HFV + GWLDSF SVM VTTVG + GDR FTGA + WL VSTLAVARAFL + LA + AR KR + R + K VLG ++ IS + F ADIDN + G + S EF IYKLK + M K ++++ D + IC FD + LD G + ITL DL +
Sbjct: 102 VALYFFVVTFC --- GFLIVHFVVKIGWLDSFCFSVMMVTTVGFGDRAFNTWLGTFLAAVWLLVSTLAVARAFLFLADARADKRNRERAKKVLGESISISISQFFAADIDNDGRLSLAEFAIYKL.225 бит (134), ожидание = 6,112e-8
Дифференциальная регуляция супероксиддисмутазы в растениях, подверженных стрессу окружающей среды, по JSTORАбстрактныйСупероксиддисмутазы (СОД) - это металлопротеины, которые катализируют дисмутацию супероксидных радикалов до перекиси водорода и кислорода.Фермент широко распространен в аэробных организмах, где он играет важную роль в защите от токсичности, опосредованной кислородными радикалами. У растений неблагоприятные условия окружающей среды часто приводят к увеличению выработки восстановленных форм кислорода, и, следовательно, предполагается, что СОД играет важную роль в устойчивости растений к стрессу. Здесь мы описываем выделение клона кДНК, кодирующего цитозольную SOD медь / цинк из Nicotiana plumbaginifolia. Используя это, вместе с ранее выделенными кДНК, кодирующими митохондриальную SOD марганца и хлоропластную железную SOD в качестве зондов в анализах гель-блоттинга РНК, мы изучили изобилие транскриптов SOD в различных стрессовых условиях: в ответ на свет, во время фотоингибирующих условий (свет в сочетании с высоким или низкие температуры), и в ответ на ксенобиотический стресс, вызванный гербицидом паракватом.Представлены доказательства того, что содержание мРНК SOD железа увеличивается всякий раз, когда возникает локализованный в хлоропласте окислительный стресс, аналогично предыдущему открытию, что SOD марганца реагирует на события, локализованные в митохондриях. Таким образом, разнообразные эффекты различных стрессовых условий на изобилие мРНК SOD могут дать представление о том, как каждое лечение влияет на различные субклеточные компартменты. Информация о журнале«Растительная клетка» выходит 19-й год.В течение трех лет после своей первой публикации он занял первое место по значимости среди основных исследовательских журналов по наукам о растениях, и с тех пор он поддерживает этот стандарт качества. The Plant Cell публикует новые исследования, имеющие особое значение в биологии растений, особенно в областях клеточной биологии, молекулярной биологии, генетики, развития и эволюции. Статьи дают новые знания, которые представляют большой интерес не только для специалистов, но и для биологов растений. Информация об издателеOxford University Press - это отделение Оксфордского университета.Издание во всем мире способствует достижению цели университета в области исследований, стипендий и образования. OUP - крупнейшая в мире университетская пресса с самым широким глобальным присутствием. В настоящее время он издает более 6000 новых публикаций в год, имеет офисы примерно в пятидесяти странах и насчитывает более 5500 сотрудников по всему миру. Он стал известен миллионам людей благодаря разнообразной издательской программе, которая включает научные работы по всем академическим дисциплинам, библии, музыку, школьные и университетские учебники, книги по бизнесу, словари и справочники, а также академические журналы. SEC.gov | Превышен порог скорости запросовЧтобы обеспечить равный доступ для всех пользователей, SEC оставляет за собой право ограничивать запросы, исходящие от необъявленных автоматизированных инструментов. Ваш запрос был идентифицирован как часть сети автоматизированных инструментов за пределами допустимой политики и будет обрабатываться до тех пор, пока не будут приняты меры по объявлению вашего трафика. Пожалуйста, объявите свой трафик, обновив свой пользовательский агент, чтобы включить в него информацию о компании. Чтобы узнать о передовых методах эффективной загрузки информации с SEC.gov, в том числе о последних документах EDGAR, посетите sec.gov/developer. Вы также можете подписаться на рассылку обновлений по электронной почте о программе открытых данных SEC, включая передовые методы, которые делают загрузку данных более эффективной, и улучшения SEC.gov, которые могут повлиять на процессы загрузки по сценарию. Для получения дополнительной информации обращайтесь по адресу [email protected]. Для получения дополнительной информации см. Политику конфиденциальности и безопасности веб-сайта SEC.Благодарим вас за интерес к Комиссии по ценным бумагам и биржам США. Идентификатор ссылки: 0.5dfd733e.1630392034.2d95eedc Дополнительная информацияПолитика безопасности в ИнтернетеИспользуя этот сайт, вы соглашаетесь на мониторинг и аудит безопасности. В целях безопасности и обеспечения того, чтобы общедоступная услуга оставалась доступной для пользователей, эта правительственная компьютерная система использует программы для мониторинга сетевого трафика для выявления несанкционированных попыток загрузки или изменения информации или иного причинения ущерба, включая попытки отказать пользователям в обслуживании. Несанкционированные попытки загрузить информацию и / или изменить информацию в любой части этого сайта строго запрещены и подлежат судебному преследованию в соответствии с Законом о компьютерном мошенничестве и злоупотреблениях 1986 года и Законом о защите национальной информационной инфраструктуры 1996 года (см. Раздел 18 USC §§ 1001 и 1030). Чтобы обеспечить хорошую работу нашего веб-сайта для всех пользователей, SEC отслеживает частоту запросов на контент SEC.gov, чтобы гарантировать, что автоматический поиск не влияет на возможность доступа других пользователей к SEC.содержание правительства. Мы оставляем за собой право блокировать IP-адреса, которые отправляют чрезмерное количество запросов. Текущие правила ограничивают пользователей до 10 запросов в секунду, независимо от количества машин, используемых для отправки запросов. Если пользователь или приложение отправляет более 10 запросов в секунду, дальнейшие запросы с IP-адреса (-ов) могут быть ограничены на короткий период. Как только количество запросов упадет ниже порогового значения на 10 минут, пользователь может возобновить доступ к контенту на SEC.губ. Эта практика SEC предназначена для ограничения чрезмерного автоматического поиска на SEC.gov и не предназначена и не ожидается, чтобы повлиять на людей, просматривающих веб-сайт SEC.gov. Обратите внимание, что эта политика может измениться, поскольку SEC управляет SEC.gov, чтобы гарантировать, что веб-сайт работает эффективно и остается доступным для всех пользователей. Примечание: Мы не предлагаем техническую поддержку для разработки или отладки процессов загрузки по сценарию. Положительный отбор, структура молекулярной рекомбинации и филогенетическая реконструкция членов семейства Tombusviridae: значение для таксономии вирусов(1)Положительный отбор, структура молекулярной рекомбинации и филогенетикареконструкция членов семейства Tombusviridae:Значение для таксономии вирусовМонсеф БулилаInstitut de l’Olivier, Sousse Ibn-khaldoun, Тунис.АбстрактныеПодробное исследование предполагаемых событий рекомбинации и частоты их эволюции во всем геноме современной известные представители семейства Tombusviridae, включающего 79 образцов, извлеченных из международных баз данных, проводился с использованием алгоритмов RECCO и RDP версии 3.31. Первая программа позволила обнаружить потенциальные сайты рекомбинации в семи из восьми родов вирусов (Aureusvirus, Avenavirus, Carmovirus, Dianthovirus, Necrovirus, Panicovirus и Tombusvirus) вторая программа дала те же результаты, за исключением рода Диантовирус.С другой стороны, оба метода не смогли обнаружить точки останова рекомбинации в геноме представители рода Machlomovirus. Кроме того, на основе точного теста нейтральности Фишера положительный отбор оказывался на гены, кодирующие белок, были обнаружены в 17 парах образцов, включающих 15 различных линий. Кроме родов Machlomovirus и Panicovirus вместе с неклассифицированными Tombusviridae, всеми другими таксономическими родами и unassignedTombusviridae - представители, прошедшие положительный отбор.Эволюционная история всего члены семейства Tombusviridae показали, что они разделились на восемь отдельных групп, соответствующих восемь родов, составляющих это семейство. Предполагаемая филогения изменила классификацию, принятую в настоящее время Международный комитет по таксономии вирусов. Была предложена реклассификация. Ключевые слова: биоинформатика, филогения, рекомбинация, положительный отбор, Tombusviridae, последовательность, таксономия. Поступила: 22 ноября 2010 г .; Принята в печать: 13 мая 2011 г. ВведениеРекомбинацияРНК является одним из основных факторов отвечает за создание новых РНК-вирусов и ретро-вирусы. Биологические механизмы рекомбинации различаются между организмами, но в широком смысле рекомбинация приводит к созданию мозаичных последовательностей, в которых эволюционная история на каждом участке может быть разной. Рекомбинация, определяемая как обмен генетической информацией между двумя нуклеотидными последовательностями, является важным процессом который влияет на биологическую эволюцию во многих различных уровни.Рекомбинация объясняет значительное количество генетическое разнообразие природных популяций и генов в целом расположены в областях генома с низким уровнем рекомбинации имеют низкий уровень полиморфизма (Posada и Крэндалл, 2001). Рекомбинация перетасовывает существующие вариация и даже создает новые варианты. Было показано, что Рекомбинация РНК позволяет обмениваться генетическими материал, не только между одними и теми же или подобными вирусами, но и между совершенно разными вирусами (Воробей и Холмс, 1999).Иногда это также допускает кроссоверы между вирусными и РНК хозяина (Грин и Эллисон, 1994; Аазиз и Теп-фер, 1999; Baroth et al. , 2000; Nagai et al. , 2003). Принимая учитывать структуру вирусных геномных молекул и расположение сайтов кроссовера, три основных типа РНК при повторной комбинации были выделены: гомологичные, аберрантные гомологичны и не гомологичны (Lai, 1992; Alejska et al. , 2001). Первые два происходят между двумя идентичными или похожие РНК (или между молекулами, показывающими локальные гомология), а в последней входят две разные молекулы.Большинство собранных данных предполагают, что рекомбинанты РНК формируются в соответствии с моделью выбора копии (Alejska et al. , 2001). Комплекс вирусной репликации запускает зарождающуюся РНК синтез цепи на одной матрице, называемой донором РНК, и затем переключается на другой шаблон, называемый акцептором РНК. Соответственно, считается, что на РНК влияют два основных фактора. рекомбинация: структура рекомбинирующих молекул и способность вирусной репликазы переключать шаблоны. Из поколения в поколение вирусные популяции развиваются различные селективные силы в разных регионах и участках, которые отображать различные функциональные ограничения.Строгий и надежный критерий обнаружения адаптивной эволюции в кодирующий белок ген является ускоренным несинонимичным ( d N, амино замена кислоты) скорость относительно синонимичной ( d S, тихая) скорость замен при соотношении скоростей = d N / d S> 1. Поскольку односторонние мутации не изменяют аминокислоту, тогда как Регенетика и молекулярная биология , 34, 4, 647-660 (2011) Авторские права © 2011, Sociedade Brasileira de Genética.Напечатано в Бразилии www.sbg.org.br Отправить корреспонденцию Moncef Boulila. Institut de l’Olivier, B.P. 14, 4061 Сусс Ибн-Халдун, Тунис. Эл. адрес: [email protected]. (2) Естьмутаций размещения, разница в их фиксации ставки обеспечивают меру избирательного давления на белок. Среди РНК-вирусов растений с положительной цепью семейство Tombusviridae включает несколько вирусов с важный экономический эффект. По данным 8-го ICTV (Международный комитет по таксономии вирусов) отчет (Fauquet et al., 2005), в семейство Tombusviridae входят следующих родов: Tombusvirus , Carmovirus , Некровирус , Диантовирус , Махломовирус , Авенавирус , Aureusvirus и Panicovirus . По данным Балтимора классификации вирусы этого семейства классифицируются как Вирусы типа IV и являются частью супергруппы лютеовирусов. (Хабили и Саймонс, 1989). РНК содержится в икосаэдрический (T = 3) капсид, состоящий из 180 единиц белок однократной оболочки размером 27-42 кДа; меры вириона 28-35 нм в диаметре, без оболочки.Все Tombusviridae имеют положительный смысл, одноцепочечные геном lin-ear, за исключением диантовирусов, чьи геном двудольный. Геном составляет примерно 4-5,4 т.п.н. в длину в зависимости от рода. 3 ’конечная точка не полиаденилированный. 5 ’конец ограничен только Кармовирус крапчатости , Клевер красный некротическая мозаика диантовирус и Махломовирус хлоротичной крапчатости кукурузы . Геном кодирует 4-6 ORF. Полимераза ORF кодирует янтарный стоп-кодон, который является местом считывания событие в ORF 1 (за исключением диантовирусов, где считывание происходит с через при сдвиге кадра), в результате получается два продукты, необходимые для репликации.Не кодируется геликазой вирусом. Процесс репликации членов семьи Tombusviridae состоит из следующих этапов: (i) вирус проникает в хозяйскую клетку, (ii) вирусная геномная РНК без оболочки и высвобождается в цитоплазму, (iii) вирусный РНК транслируется с образованием двух белков, необходимых для Синтез РНК (репликация и транскрипция), (iv) a комплементарная оцРНК с отрицательным смыслом синтезируется с использованием геномная РНК в качестве матрицы, (v) новая геномная РНК синтезируется с использованием отрицательно-смысловой РНК в качестве матрицы, (vi) РНК-зависимая РНК-полимераза (RdRp) распознает внутренние субгеномные промоторы по негативному РНК для транскрипции 3’-концевых субгеномных РНК который будет генерировать капсид и белок движения, (vii) образуются новые вирусные частицы (White and Nagy, 2004). Основной целью данной работы было определение и охарактеризовать механизмы эволюции вирусов Tombusviridae на основании появления предполагаемых рекомбинационные события и положительный отбор в их полном объеме геном. Это было достигнуто путем анализа 79 образцов. получено в GenBank. В результате мы предлагаем переклассификация в соответствии с их предсказанной эволюционной историей. Материалы и методыПоследовательности всего генома 79 образцов каталогизированные в GenBank были использованы в данном исследовании (Таблица 1). Нуклеотидные последовательности выравнивали с использованием программы CLUSTALW 2.0.9 и CLUSTALX 2.0.9 (Larkin и др. , 2007) с конфигурацией по умолчанию. Их филогенетические отношения были определены с Алгоритм максимального правдоподобия (ML), включенный в Программа MEGA версии 5 (Tamura et al. , 2011) под предположение моделей замещения, предложенных Джуксом и Кантор (1969) (JC), Hasegawa et al. (1985) (HKY85) и Тамура и Ней (1993) (TN93).Анализ начальной загрузки с помощью Было выполнено 500 повторов для оценки устойчивости ветви. Используя программу MEGA4.1b (Кумар и др. , 2008), положительный отбор был сделан методом подсчета описанный Nei и Gojobori (1986), а затем Suzuki и Годжобори (1999). Согласно этому методу Использовано филогенетическое дерево анализируемых последовательностей. Для метод парсимонии, общее количество синонимов ( c S) и несинонимичные ( c N) замены, а также среднее номера синонимичных ( s S) и несинонимичных ( s N) сайтов на кодон по филогенетическому дереву для каждого кодонного сайта были рассчитаны в соответствии с максимальной экономией принцип (Fitch, 1971; Hartigan, 1973).Нулевая гипотеза селективная нейтральность ( r S = r Norw = 1) была проверена для каждого сайта путем вычисления вероятности ( p ) получения наблюдаемого или более смещенные значения для c Sand c N, которые предполагались равными следовать биномиальному распределению с вероятностями возникновение синонимичных и несинонимичных замен задается как с S / ( с S + с N) и с N / ( с S + с N), соответственно.Положительный выбор делается при p <0,05 и c N / s N> c S / s S (Су-дзуки, 2006). Возможные события рекомбинации между расходящимися нуклеотидные последовательности исследовали с помощью двух программ: RDP v3.31b (Martin и др. , 2005b) и RECCO (Maydt и Ленгауэр, 2006). RDP включает в себя несколько опубликованных повторное объединение методов обнаружения в единый набор инструментов: RDP (Мартин и Рыбицки, 2000), GENECONV (Падидам и др., 1999), BOOTSCAN (Мартин и др. , 2005a), MAXCHI (Смит, 1992), CHIMAERA (Посада и Cran-dall, 2001), SISCAN (Gibbs и др. , 2000) и 3SEQ (Boni и др. , 2007). Во всех случаях использовались параметры по умолчанию. Только события, предсказанные более чем половиной методов, являются считается значительным. Алгоритм разработан и описанный Мейдтом и Ленгауэром (2006) как быстрый, простой и чувствительный метод обнаружения рекомбинации в набор последовательностей и расположение предполагаемой рекомбинации точки останова основаны на минимизации затрат.Этот метод имеет только два настраиваемых параметра, рекомбинация и мутация Стоимость. На практике единственным рассматриваемым параметром является a, представляет стоимость мутации по сравнению с рекомбинацией. При изменении от 0 до 1 стоимость мутации взвешивается. увеличивается, а стоимость рекомбинации, взвешенная на 1 -увеличивается. Другими словами, параметр управляет неоднозначность между мутацией и рекомбинацией. (3)эволюция Tombusviridae 649 Таблица 1 - Текущий таксономический статус членов семейства Tombusviridae, включенных в исследование, и их инвентарные номера. Род (жирным шрифтом), неклассифицированный и неназначенные вирусы Virus / Isolate Регистрационный номер GenBank Aureusvirus Вирус пятнистости листьев огурца (CLSV) NC_007816 Вирус пятнистости листьев огурца / Канада (CLSV / Canada) EU127904 Латентный вирус Pothos / гороха голубя (PoLV / горох голубя) NC_000939 Вирус мозаики хлоротичных полос Johnsongrass / Иран (JCSMV / Iran) NC_005287 Вирус мозаики белой линии кукурузы / США (MaWLMV / USA) NC_009533 Авенавирус Вирус хлоротической задержки овса (OCSV) NC_003633 Кармовирус Вирус хлоротических пятен кардамина (CCFV) NC_001600 Вирус крапчатости гвоздики / Китай (CarMoV / China) NC_001265 Вирус крапчатости гвоздики (CarMoV) X02986 Вирус крапчатости гвоздики / индийский (CarMoV / Indian) AJ811998 Вирус крапчатости коровьего гороха (CPMoV) NC_003535 Вирус хлоротической кольцевой пятнистости гибискуса (HCRSV) NC_003608 Вирус хлоротической кольцевой пятнистости гибискуса.Tw (HCRSV.Tw) DQ392986 Вирус некротического кольца радужки японской (JINRV) NC_002187 Вирус некротической пятнистости дыни / Ямагути (MeNSV / Ямагути) AB250687 Вирус некротической пятна дыни / Nagasaki (MeNSV / Nagasaki) AB250686 Вирус некротической пятнистости дыни / Kochi (MeNSV / Kochi) AB250685 Вирус некротической пятна дыни / Chiba (MeNSV / Chiba) AB250684 Вирус некротической пятна дыни / Тоттори (MeNSV / Tottori) AB232925 Вирус некротической пятнистости дыни / Kochi2 (MeNSV / Kochi2) AB232926 Вирус некротической пятнистости дыни (MeNSV) NC_001504 Вирус некротической пятнистости дыни / MNSV-ISR (MeNSV / MNSV-ISR) DQ922807 Вирус некротической пятнистости дыни / MNSV-Al (MeNSV / MNSV-Al) DQ339157 Вирус некротической пятна дыни / MNSV264 (MeNSV / MNSV264) AY330700 Вирус некротической пятна дыни / nK (MeNSV / nK) AB044292 Вирус некротической пятна дыни / NH (MeNSV / NH) AB044291 Вирус некротической пятнистости дыни / Malfa5 (MeNSV / Malfa5) AY122286 Вирус некроза стеблей гороха / Япония (PSNV / Japan) NC_004995 Вирус разрушения цветков пеларгонии / MZ10 (PFBV / MZ10) NC_005286 Вирус разрушения цветков пеларгонии / SP18 (PFBV / SP18) DQ256073 Вирус кактуса сагуаро (SCV) NC_001780 Вирус морщинистости репы (TCV) NC_003821 Вирус морщинистости репы / Великобритания (TCV / UK) AY312063 Неклассифицированный вирус надрыва цветков Ангелония / Флорида (AFBV / Флорида) NC_007733 Кармовирусы Вирус желтой крапчатой мозаики сои / MS1-USA (SYMoMV / MS1-USA) FJ707484 Вирус желтой крапчатой мозаики сои / Южная Корея (SYMoMV / s.Корея) NC_011643 Диантовирус РНК 1 вируса кольцевой пятнистости гвоздики (CarRSV-RNA 1) NC_003530 РНК 2 вируса кольцевой пятнистости гвоздики (CarRSV-RNA 2) NC_003531 РНК 1 вируса некротической мозаики красного клевера (RCNMV-RNA 1) NC_003756 РНК 2 вируса некротической мозаики красного клевера (RCNMV-RNA 2) NC_003775 РНК вируса некротической мозаики красного клевера 1 / Can (RCNMV-RNA 1 / Can) AB034916 РНК 2 / Can вируса некротической мозаики красного клевера (RCNMV-RNA 2 / Can) AB034917 РНК вируса некротической мозаики сладкого клевера 1/59 (SCNMV-RNA 1/59) NC_003806 (4)РезультатСобытия рекомбинации во время Tombusviridae эволюция Проверка результатов программы RECCO учитывая возникновение рекомбинационных событий в полном геном семейства Tombusviridae выявил, что три из пяти вирусов aureus были предположительно рекомбинантами (PoLV.Pigeonpea, JCSMV, Иран, MaWLMV, США). В напротив, CLSV (неизвестный изолят) и CLSV. Канада не показывают любой рекомбинантный сигнал (Таблица 2). Внутри рода Aureusvirus , наиболее часто рекомбинирующим вирусом был PoLV.Pigeonpea (33 предполагаемых сайта рекомбинации), тогда как только 28 возможных сигналов рекомбинации были обнаружены в геноме вирусов JCSMV.Iran и MaWLMV.USA. Точно так же единственный представитель род Avenavirus (OCSV) был потенциальным рекомбинантным со 175 предполагаемыми сайтами.Пакет RDP подтвердил эти результаты для обоих родов. Среди кармовирусов 14 из Возможными рекомбинантами были 30 членов. В соответствии с RECCO, наиболее часто рекомбинирующий вирус был JINRSV со 134 предполагаемыми событиями, а MeNSV.Nagasaki 650 Boulila Род (жирным шрифтом), неклассифицированный и неназначенные вирусы Virus / Isolate Регистрационный номер GenBank Неклассифицированный диантовирус X РНК 1 вируса риса (RVX-RNA 1) AB033715 Махломовирус Вирус хлоротической крапчатости кукурузы (MCMoV) NC_003627 Вирус хлоротической крапчатости кукурузы / Небраска (MCMoV / Небраска) EU358605 Некровирус Вирус черного ожога свеклы (BBSV) NC_004452 Вирус черного ожога свеклы / Val25-Иран (BBSV / Val25-Iran) EU545828 Вирус ожога черной свеклы / CO-USA (BBSV / CO-USA) EF153268 Вирус черного ожога свеклы / Синьцзян (BBSV / Синьцзян) AY626780 Вирус белой полосы лука-порея (LWSV) NC_001822 Оливковый латентный вирус 1 / Citrus (OLV-1 / Citrus) NC_001721 Оливковый латентный вирус 1 / GM6-Portugal (OLV-1 / GM6-Portugal) DQ083996 Вирус некроза табака A / FM1B (TNV-A / FM1B) NC_001777 Вирус некроза табака A / C (TNV-A / C) AY546104 Вирус некроза табака D / венгерский (TNV-D / венгерский) NC_003487 Вирус некроза табака D / Rhotamsted (TNV-D / Rhotamsted) D00942 Некровирус, неклассифицированный Вирус мягкой мозаики оливы / GP-POrtugal (OMMV / GP-Portugal) NC_006939 Паниковирус Вирус мозаики паникум (PMV) NC_002598 Tombusvirus Вирус пятнистой морщинистости артишока / Бари (AMoCV / Bari) NC_001339 Вирус кольцевой пятнистости итальянской гвоздики (CarIRSV) NC_003500 Болгарский латентный вирус огурца (CBLV) NC_004725 Вирус некроза огурца (CNV) NC_001469 Вирус кольцевой пятнистости Cymbidium (CymRSV) NC_003532 Алжирский латентный вирус виноградной лозы / нипплфрут (GALV / нипплфрут) NC_011535 Латентный вирус груши (PeLV) NC_004723 Вирус кустистого трюка томатов / Statice (TBSV / Statice) AJ249740 Tomato bushy stunt virus / Nipplefruit (TBSV / Nipplefruit) AY579432 Вирус кустистого каскадного роста томатов / Nipplefruit (TBSV / Nipplefruit) Вирус кустистого трюка помидоров / перец (TBSV / Pepper) U80935 Tomato bushy stunt virus / Cherry (TBSV / Cherry) M21958 - Вирус кустистого каскадерства томатов / вишня (TBSV / Cherry) Неклассифицированные вирусы томбуса Вирус некроза лизиантуса / л (LNV / L) NC_007983 Вирус некроза лизиантуса / Zantedeschia (LNV / Zantedeschia) AM711119 Вирус некротической пятна пеларгонии (PNSV) NC_005285 Неназначенный Tombusviridae Вирус некротической полосы кукурузы (MaNSV) Вирус паттерна линий пеларгонии / PV-0193 (PLPV / PV-0193) NC_007729 NC_007017 Неклассифицированный Tombusviridae Вирус волчаночного очищения Нутка / Аляска (NLVCV / Аляска) Вирус хлоротического кольца пеларгонии / GR 57 (PCRPV / GR 57) (5)Томбусвириды эволюция 651 Таблица 2 - Определение предполагаемых предполагаемых событий рекомбинации и их частоты по последовательностям всего генома вирусов aureus, одного авенавируса и кармовирусов.Показан алгоритм RDP v3.31b. что сообщаются только события, поддерживаемые более чем половиной различных методов. Нумерация нуклеотидов соответствует выровненным последовательностям. Сокращения: NRS: -количество сайтов рекомбинации, GIRE: -геномный интервал событий рекомбинации (диапазон последовательностей в вирусном геноме, где были предсказаны события рекомбинации). Рекомбинация определена RECCO Рекомбинация определена RDP v3.31b Virus.isolate Длина точки останова Геномная позиция самой длинной точки останова (размер в нуклеотидах) Предполагаемый родитель (Major x Minor) NRS GIRE (нуклеотид) 1 остаток 2 остатка 3 остатка> 3 остатка PoLV.Голубиный горох 33 607-5320 10 8 3 12 2148-2184 (37) OCSV x MeNSV / MNSV-264 РНК CarRSV 1 x MeNSV-Tottori CarRSV РНК 1 x MeNSV-Kochi 2 JCSMV.Iran 28 2457-5288 11 6 3 8 3790-3802 (13) РНК CarRSV 1 x MeNSV-Tottori CarRSV РНК 1 x MeNSV-Kochi 2 MaWLMV.USA 28 3212-4043 18 3 1 6 3836-3853 (18) OCSV x MeNSV / MNSV-264 РНК CarRSV 1 x MeNSV-Tottori OCSV 175 766-5052 65 33 23 54 1295-1306 (12) 5036-5047 (12) TCV.Великобритания x MaWLMV.USA CCFV 20 894-4681 7 2 3 8 1896-1908 (13) / CarMoV 5 1349-2124 0 0 1 4 1986-2045 (60) / CPMoV 14 1316-3143 4 6 2 2 2850-2854 (5) / ОИЯИ 134 689-5098 58 30 20 26 1736-1760 (25) / MeNSV.Nagasaki 2 623-4892 0 0 0 2 4847-4892 (46) / МеНСВ.Кочи 4 1843-4694 1 0 0 3 3866-3907 (42) / MeNSV.MNSV-Al 3 619-4908 0 0 0 3 4864-4908 (45) / МНСВ.МНСВ-264 5 4940-5131 3 0 2 0 4952-4954 (3) 5129-5131 (3) / MeNSV.NK 2 615-4908 0 1 0 1 615-874 (60) SYMoMV / MS1-USA x TBSV.Statice MeNSV.NH 5 615-4983 1 1 0 3 2070-2151 (82) YMoMV / MS1-USA x TBSV.Statice MeNSV.Malfa5 13 677-4831 0 0 2 11 1736-1796 (61) / ПСНВ. Япония 80 645-5174 34 15 9 22 4225-4266 (42) / SCV 61665-4506 28 6 13 14 1887-1898 (12) 2761-2772 (12) / (6)и MeNSV.NK имели наименьшее число рекомбинаций. сайты (два предполагаемых сайта). Алгоритм RDP v3.31 подтвердил наличие возможных событий рекомбинации только для присоединений MeNSV.NK и MeNSV.NH. Рекомбинационные исследования рода Dianthovirus на основе Анализ RECCO показал, что только 80% из них были возможные рекомбинанты (CarRSV-RNA 1 и 2, RCNMV-RNA 1 и 2, RCNMV.Can. РНК 1 и 2, SCNMV.59.RNA 1, и RVX.RNA 1). напротив, SCNMV.59.RNA 2 и SCNMV.38.RNA не обнаружил предполагаемой рекомбинации. сигналы. Однако пакет RDP не предсказал рекомбинация у диантовирусов (таблица 3). Хотя большинство часто рекомбинирующим некровирусом был RVX (166 предполагаемых сайты), RCNMV.РНК 2 имела только два предполагаемых сайта. На основании по анализу RECCO 50% некровирусов (BBSV.Val25.Iran, LWSV, TNV-A.FMB, TNV-A.C, TNV-D.Hungarian и OMMV-GP.Portugal,) были возможные рекомбинанты. И наоборот, BBSV, BBSV.CO.USA, BBSV.Xinjiang, OLV-1.Citrus, OLV-1.GM6.Portugal, и TNV-D.Rhotamsted не были возможными рекомбинантами. Эти результаты совпадали с результатами, полученными с помощью Пакет RDP. Хотя наиболее часто рекомбинирующие vi-rus был LWSV (39 сайтов), TNV.A.C рекомбинирован в два места.По поводу единственного представителя рода Panicovirus (PMV), результаты, полученные двумя методы (RECCO и RDP v3.31b) несовместимы. Действительно, с RECCO было обнаружено 108 возможных сайтов, в то время как сигналы рекомбинации были обнаружены с помощью пакета RDP. Похожая ситуация наблюдалась и с новым предложенные кармовирусы (NLVCV.Alaska, PCRPV.GR 57, ПЛПВ.ПВ-0193). Согласно анализу RECCO, хотя NLVCV.Alaska была наиболее часто рекомбинирующей vi-rus (65 сайтов), PLPV.PV-0193 перекомпонован только в 38 сайты (таблица 4). Относительно представителей рода Tombusvirus , между двумя методы, показывающие, что 80% проанализированных образцов были предполагаемыми рекомбинантами. В то время как у CBLV был самый высокий количество предполагаемых сигналов рекомбинации (67 сайтов), У TBSV.Cherry было всего два сайта рекомбинации. Более того, следует отметить, что два представителя рода Махломовирус (MCMoV и MCMoV.Nebraska) были не рекомбинанты по оценке двумя методами анализ используется в этом исследовании.В поисках рекомбинации частота в геноме Tombusviridae, две трети aureusviruses (JCSMV.Iran и MaWLMV.USA) показали, что в большинстве случаев длина их точки останова была одиночный остаток. Напротив, длина точки останова большинства предполагаемые сайты рекомбинации PoLV. от 3 до 37 нуклеотидов (таблица 2). Так же длина точки излома основных сайтов рекомбинации одиночного представитель рода Avenavirus (OCSV) состоял из одиночный остаток.Примерно у 50% представителей рода Carmovirus , длина наиболее детектируемого сайты рекомбинации представляли собой один остаток. В отличие от этого, интервал точки останова для остальных участников превышен три остатка размером до 82 остатков (MeNSV.NH). В 62% исследованных диантовирусов длина точки останова превышала три нуклеотида, достигая 100 остатков (CarRSV.RNA 1) (таблица 3). в некровирусов, интервал распределения точек останова был аналогичный i.е. , 50% контрольных точек состояли из одного остатка, в то время как оставшиеся точки останова были между тремя и 77 нуклеотиды. Для единственного представителя рода Panicovirus (PMV), большинство сайтов рекомбинации имели точку останова длина одиночного остатка (45) (таблица 3). Для tombusviruses, 75% показали, что длина точки останова превышает три остатка до 161 нуклеотида (AMoCV.Bari) (Таблица 4). Анализ нуклеотидной последовательности Оценка максимального совокупного правдоподобия Паттерн нуклеотидных замен был составлен с использованием MEGA4.1б программа. Результаты для Tombusviridae показали, что ставки различных переходных замен варьировались от 3,18 до 14,61, а поперечных замен варьировали от 6,6 до 8,57. Нуклеотид частоты: 0,269 (A), 0,258 (T / U), 0,207 (C) и 0,266 (G). Соотношение скорости перехода / трансверсии составляло k 1 = 1,705. (пурины) и k 2 = 0,482 (пиримидины). Общий переходное смещение / поперечное смещение составляло R = 0,547, где R = [AG k 1+ TC k 2] / [(A + G) (T + C)].Всего было 1218 позиций в окончательном наборе данных. Во всех этих анализах кодон Включенные позиции: первая + вторая + третья + некодирование. Все позиции, содержащие пробелы и недостающие данные, были исключены из набора данных (возможность полного удаления). Программа MEGA4.1b также включает Тест Таджи-ма на нейтралитет. Цель этого теста - определить последовательности, которые не соответствуют модели нейтральной теории при равновесие между мутацией и генетическим дрейфом. Тест Таджимы сравнивает стандартизированную меру общего количества сайты сегрегации (полиморфные сайты ДНК) в взятая ДНК и среднее количество мутаций между пары в выборке.Тадзима D был определен ( D = 5,280926). Положительный выбор К высокой генетической стабильности вирусов можно отнести к отрицательному или очищающему отбору для поддержания функционального целостность вирусного генома. Степень отрицательного селекции в генах, или степень функционального ограничения для поддержание кодируемой белковой последовательности, может быть оценивается, как упоминалось выше, соотношением между нуклеотидное разнообразие несинонимичных и синонимичных положения ( d N / d S).Для большинства кодирующих генов соотношение d N / d <1 что согласуется с отрицательной селекцией против белка изменение. Напротив, d N / d Sratio> 1 может быть показателем что адаптивный или положительный отбор движет геном дивергенция. В этом исследовании парные сравнения всех отобранных образцы показали, что ни один из представителей родов Махломовирус и Паниковирус, и неклассифицированные (7)Томбусвириды эволюция 653 Таблица 3 - Определение предполагаемых предполагаемых событий рекомбинации и их частоты по последовательностям некровирусов, одного паниковируса и диантовирусов.Алгоритм RDP v3.31b показал, что только события сообщается, что поддерживается более чем половиной различных методов. Нумерация нуклеотидов соответствует выровненным последовательностям. Сокращения: NRS: -количество сайтов рекомбинации, GIRE: -геномный интервал события повторной комбинации (диапазон последовательностей в вирусном геноме, где были предсказаны события повторной комбинации). Рекомбинация определена RECCO Рекомбинация определена RDP v3.31b Virus.isolate Длина точки останова Геномная позиция самой длинной точки останова (размер в нуклеотидах) Предполагаемый родитель (Major x Minor) NRS GIRE (нуклеотид) 1 остаток 2 остатка 3 остатка> 3 остатка CarRSV-РНК 1 19 709-4561 7 3 2 7 2581-2680 (100) / CarRSV-РНК 2 3 1826-2811 0 0 0 3 1826-1835 (10) 2802-2811 (10) / RCNMV-РНК 1 8 1297-4754 0 1 2 5 1479-1533 (55) / RCNMV-РНК 2 2 2696-2812 1 0 0 1 2803-2812 (10) / RCNMV-RNA1.Банка 7 720-4218 2 0 0 5 1556-1644 (89) / RCNMV-РНК 2. Может 10 1730-2827 3 1 2 4 1730-1765 (36) / SCNMV-РНК 1,59 12 858-4443 2 2 1 7 3126-3193 (68) / RVX-РНК 1 166439-5107 65 39 19 43 840-855 (16) / BBSV-Val25.Iran 4 800-4881 2 0 0 2 1435-1468 (34) MeNSV.NH x LNV.Zantedeschia MeNSV.Malfa5 x MaNSV SCV x CNV MeNSV.Yamaguchi x CNV LWSV 39 1027-4853 17 9 5 8 4846-4853 (8) MeNSV.NH x LNV.Zantedeschia SCV x CNV ТНВ-А.FM1B 3 2419-2789 3 0 2 1 2419-2496 (77) OLV1.GM6-Portugal x BBSV.Xinjiang TNV-A.C 2 2275-2619 0 0 0 2 2275-2619 (10) OLV1.GM6-Portugal x BBSV.Xinjiang LWSV x TNV.D. Ротамстед TNV-D. Венгрия 4 2323-4287 2 0 0 2 3959-3964 (6) MeNSV.NH x LNV.Zantedeschia MeNSV.Malfa5 x MaNSV BBSV x OMMV.GP.Португалия SCV x CNV OMMV-GP. Португалия 20 935-4857 6 3 4 7 2116-2145 (30) TNV.A.C x TNV.D. Венгрия TNV.A.C x TNV.Д. Ротамстед TNV.D.Rhotamsted x TNV.A.FM1B OLV1.GM6-Portugal x TNV.D. Венгерский (8)654 Boulila Таблица 4 - Определение предполагаемых предполагаемых событий рекомбинации и их частоты в последовательностях вирусов tombus и предполагаемых новых членов рода Carmovirus . Показан алгоритм RDP v3.31b. что сообщаются только события, поддерживаемые более чем половиной различных методов. Нумерация нуклеотидов соответствует выровненным последовательностям.Сокращения: NRS: -количество сайтов рекомбинации, GIRE: -геномный интервал событий рекомбинации (диапазон последовательностей в вирусном геноме, где были предсказаны события рекомбинации). Рекомбинация определена RECCO Рекомбинация определена RDP v3.31b Virus.isolate Длина точки останова Геномная позиция самой длинной точки останова (размер в нуклеотидах) Предполагаемый родитель (Major x Minor) NRS GIRE (nt) 1 остаток 2 остатка 3 остатка> 3 остатка AMoCV.Бари 27 1074-4774 2 4 3 18 4614-4774 (161) CymRSV x CBLV GALV.nipplefruit x MeNSV.Yamaguchi CarIRSV 28 1998-5345 9 5 8 6 4775-4842 (68) TBSV.pepper x PNSV TBSV.Statice x MeNSV.Tottori AMoCV.Bari x PNSV LNV.Zantedeschia x MaNSV SCV x CBLV ПНСВ х ТБСВ.Перец CBLV x MeNSV.MNSV-ISR LNV.Zantedeschia x TBSV.Statice CBLV 67 343-5390 16 17 10 24 1804-1836 (33) TBSV.Pepper x JCSMV.Iran CarRSV.RNA 1 x MeNSV.Tottori GALV.nipplefruit x MaWLMV.USA PeLV x MaWLMV.USA LNV.Zantedeschia x MaNSV CymRSV x MaNSV CNV 34 417-5280 8 2 6 18 2681-2714 (34) CymRSV x MeNSV ТБСВ.Перец х ПНСВ CymRSV x SCV LNV.L x TBSV.Перец CymRSV x CBLV TBSV.Statice x LNV.L CymRSV 44 675-5066 9 9 4 22 1719-1767 (49) PNSV x TBSV.Перец CarIRSV x CLSV.Израиль TBSV.Statice x TCV.UK CNV x CLSV, Канада CNV x TCV.UK MeNSV.MNSV.ISR x PeLV GALV.сосок 45 369-5299 7 4 11 23 4481-4532 (52) TBSV.Statice x CNV TBSV.nipplefruit x MaNSV TBSV.nipplefruit x TBSV.Pepper TBSV.Statice x PeLV CymRSV x CBLV CBLV x PNSV CymRSV x PeLV AMoCV.Bari x TBSN.Перец (9)Томбусвириды эволюция 655 Рекомбинация определена RECCO Рекомбинация определена RDP v3.31b Virus.isolate Длина точки останова Геномная позиция самой длинной точки останова (размер в нуклеотидах) Предполагаемый родитель (Major x Minor) NRS GIRE (nt) 1 остаток 2 остатка 3 остатка> 3 остатка ТБСВ.statice 11 379-2743 1 0 1 9 1902-1931 (30) GALV.nipplefruit x LNV.Zantedeschia CymRSV x CBLV CymRSV x PeLV TBSV.nipplefruit x TBSV.Pepper TBSV.nipplefruit 7618-1931 0 0 1 6 1550-1651 (102) GALV.nipplefruit x LNV.Zantedeschia AMoCV.Bari x GALV. Нипплефрут TBSV.pepper 7 2167-4890 0 0 1 5 4852-4890 (39) CymRSV x CBLV PeLV x AMoCV.Bari GALV.nipplefruit x LNV.Zantedeschia ТБСВ. Вишня 2 5324-5345 0 0 2 0 5324-5326 (3) 5343-5345 (3) CymRSV x CBLV CarIRSV x AMoCV.Бари CymRSV x PeLV GALV.nipplefruit x LNV.Zantedeschia MaNSV 54 547-5334 29 4 7 14 1932-1955 (24) GALV.nipplefruit x MaWLMV.USA PeLV x MaWLMV.USA ПНСВ 24 2025-5032 7 2 3 12 5009-5032 (23) TBSV.Statice x MeNSV.Tottori CNV x CLSV, Канада AMoCV.Bari x TBSV.Черри TBSV.nipplefruit x PeLV LNV.Zantedeschia x MaNSV TBSV.Statice x CNV CBLV x MeNSV.MNSV.ISR LNV.Zantedeschia x TBSV.Statice NLVCV.Аляска 65 771-5301 28 13 7 17 771-782 (12) / PCRPV.GR 57 45 1109-5104 11 17 4 13 2307-2316 (20) / ПЛПВ.ПВ-0193 38 991-4908 20 7 2 9 2813-2821 (9) / (10)Tombusviridae прошли положительный отбор. На Напротив, роды Aureusvirus (JCMSV, Иран), Avenavirus (OCSV), Кармовирус (CarMoV.China, CarMoV.Indian), Диантовирус (CarRSV-RNA 2, RCNMV-RNA 2, SCNMV-РНК 2.59, SCNMV-РНК 2.38), Некровирус (BBSV, BBSV.Val25.Iran), Томбувирус (GALV.nip-plefruit, PeLV, TBSV.Statice, PNSV) вместе с неподписанные Tombusviridae (PLPV.PV.0193) находились под положительный отбор (таблица 5). Стоит отметить, что в вирусов с сегментированным геномом положительный отбор был обнаруживается только в РНК 2, что позволяет предположить, что, вероятно, произошли события перераспределения. Все эти результаты были полученный путем проверки нейтральности в парах последовательностей с помощью Фишера Точный тест.Вероятность отклонения нулевой гипотезы строгого нейтралитета ( d N = d S) в пользу положительного отбора для каждой пары последовательностей определяли. Значения p меньше более 0,05 считались значимыми на уровне 5%. В дисперсия разницы ( d N- d S) была вычислена с использованием метод начальной загрузки (500 повторов). Все анализы были сделаны используя метод Nei-Gojobori, включенный в MEGA программа. Все позиции, содержащие пробелы и недостающие данные были исключены из набора данных (вариант полного удаления).Окончательный набор данных включал в себя 234 позиции. Филогенетические отношения Филогенетические отношения между членами семейство Tombusviridae, исходя из последовательностей их полный геном, были выведены с использованием максимума Алгоритм правдоподобия в предположении трех моделей замена (JC, HKY85, TN93). Топологии построенных дерева были идентичными. Предполагаемая филогения показали, что каждый таксономический род в семействе Tombusviridae явно составляли однородную группу. отличается от других.Однако результаты, полученные в это исследование показало несколько различий с точки зрения вируса видовой состав в пределах каждого таксономического рода сравнивается согласно текущей классификации, принятой ICTV. По факту, три вируса, которые ICTV считает неназначенными (PLPV.PV-0193) и неклассифицированные Tombusviridae (NLVCV.Alaska, PCRPV.GR 57) показали близкую филогенетическое родство с известными представителями рода Кармовирус . Более того, вирусы, относящиеся к этому роду были разделены на две отдельные подгруппы.Первое подгруппа включала вирусы: TCV, CCFV, JINRV, HCRSV, PLPV, PCRPV, NLVCV, SCV, AFBV, PFBV, CPMoV, SYMoMV и CarMoV, а вторая подгруппа зараженные вирусы: MeNSV и PSNV. Кроме того, это было предложил, чтобы род Necrovirus состоял из две отдельные подгруппы, названные предварительная подгруппа I (BBSV, TNV.D, LWSV) и предварительная подгруппа II (OMMV, ТНВ.А, ОЛВ-1) (рисунок 1). Следует отметить, что здесь OMMV является неотъемлемой частью подгруппы I, а не неклассифицированный Некровирус .В отличие от вируса рода Aureusvirus окруженные члены, которые эволюционировали в однородную манер: CLSV, PoLV, MaWLMV и JCSMV. Аналогичным образом следующие представители рода Tombusvirus также сформировали когерентный ансамбль: MaNSV, CBLV, LNV.L, LNV.Zan-tedeschia, PeLV, CNV, CymRSV, AMoCV, TBSV.Stat-ice, TBSV.Nipplefruit, TBSV.Pepper, TBSV.Cherry, GALV, PNSV и CarIRSV. Их эволюционная история 656 Boulila Таблица 5 - Образцы пар в семействе Tombusviridae при положительном отборе с вероятностью, определенной на уровне 5%, на основе точного критерия Фишера Нейтральность и расчетная d N- d S. Пары образцов Вирус.изолированные пары Значение p на уровне 5% d N- d S NC_007017 / NC_003775 PLPV / PV-0193 / RCNMV-RNA 2 0,028 0,923 AB034917 / NC_003633 RCNMV-РНК 2. Can / OCSV 0,016 2,325 NC_003531 / NC_003633 CarRSV-RNA 2 / OCSV 0,039 1,820 S46027 / NC_003633 SCNMV-РНК 2,38 / OCSV 0,041 1,847 NC_003775 / NC_003633 PLPV.PV-0193 / OCSV 0,048 1,975 NC_003807 / NC_003633 SCNMV-RNA 2.59 / OCSV 0.033 1,999 AB034917 / NC_001265 RCNMV-RNA 2. Can / CarMoV.China 0,034 1,620 AB034917 / AJ249740 RCNMV-RNA 2. Can / TBSV.Statice 0,029 1,921 NC_003531 / NC_005287 CarRSV-RNA 2 / JCSMV. Иран 0,037 1,790 S46027 / EU545828 SCNMV-РНК 2.38 / BBSV.Val25-Иран 0,045 1,682 AB034917 / AJ811998 RCNMV-RNA 2 / CarMoV. Инд. 0,031 1,679 AB034917 / NC_011535 RCNMV-RNA 2 / GALV. Плод соска 0,041 1,933 NC_003775 / NC_011535 PLPV / PV-0193 / GALV.nipplefruit 0,033 1,921 NC_003807 / NC_005285 SCNMV-RNA 2.59 / ПНСВ 0,049 1,773 S46027 / NC_004723 SCNMV-РНК 2,38 / PeLV 0,047 1,735 S46027 / NC_004452 SCNMV-РНК 2,38 / BBSV 0,043 1,749 (11)Эволюция Tombusviridae 657 (12)изменила существующую классификацию, принятую ICTV. с 2009 года. Фактически по этой классификации MaNSV считался неприсоединившимся Tombusviridae, тогда как LNV и PNSV были включены в неклассифицированные Группа Tombusvirus . Относительно рода Dianthovirus , который явно не был монофилетическим, образец кластеризации показали две отдельные клады, представляющие их РНК 1 и 2, как показано на рисунке 1.Первоначально RVX был считается неклассифицированным вирусом в пределах рода Dianthovirus . ОбсуждениеЭто исследование подтвердило предсказание предполагаемого повторная комбинация событий в геноме нескольких членов семейство Tombusviridae и продемонстрировали, что вирусы-гробницы и вирусы-гномы являются высоко рекомбинантными по сравнению к вирусам других родов. Для этого два были выбраны методы (RECCO и RDP v.3.31b), основанные на факт, что они подходят для мозаичной структуры vi-ruses, как сообщалось в предыдущих работах (Boulila, 2009; 2010).В этом исследовании с использованием алгоритма RECCO было продемонстрировали, что вирусы следующих родов содержали в своем геноме предполагаемые сигналы рекомбинации: Aureusvirus , Avenavirus , Carmovirus , Dianthovirus , Necrovirus , Panicovirus и Tombusvirus . Эти результаты были в хорошем согласии с полученными RDP пакет, кроме представителей рода Diantovirus .Оба методы, два представителя рода Machlomovirus (MCMoV, MCMoV.Nebraska) оказались нерекомбинантный. Как выяснил RECCO, наиболее часто рекомбинирующими вирусами были: OCSV, RVX.RNA 1, JINRSV, и PMV с предполагаемыми 175, 166, 134 и 108 сайты рекомбинации соответственно. Все эти сигналы рекомбинации были составлены одним остатком. MeNSV.Nagasaki, MeNSV.NK, RCNMV.RNA 2, TNV-A.C и ТБСВ.Черри (2 участка), МеНСВ.MNSV-Al, CarRSV-РНК 2, TNV-A.FM1B (3 сайта), MeNSV.Kochi, BBSV-Val25.Iran и TNV-D.Hungarian (4 сайта), CarMoV, MeNSV.MNSV-264 и MeNSV.NH (5 сайтов), RCNMV.RNA 1.Can, TBSV.nipplefruit и TBSV.pep-per (7 сайтов) и RCNMV.RNA 1 (8 сайтов) показали самая низкая частота точек останова рекомбинации. Наоборот, большинство этих точек останова имели интервал, превышающий три нуклеотиды. Кроме того, это исследование показало, что рекомбинация может происходить между вирусами, принадлежащими к разным роды.Например: Овсяный хлоротический трюк, авенавирус . (OCSV) и Кармовирус некротической точки дыни (MeNSV) может вызвать Pothos latent aureusvirus (PoLV). Аналогично, сам OCSV может быть результатом рекомбинации между Кармовирус морщинистой репы (TCV) и Кукуруза белая линия mosaic aureusvirus (MaWLMV) (таблица 2). Казалось бы, эти вирусы могли содержать часть своих последовательностей. особенно в генах, кодирующих белок оболочки друг друга.Такое событие широко изучалось для Некроз огурца tombusvirus (CNV) и Некротическая пятнистость дыни, кармовирус (MeNSV) (Ривьер и Рошон, 1990). С другой стороны, исследования селективных давление, действующее на экспрессию белков вирусных генов, привело к выявление положительного отбора в 17 парах образцов включает 15 различных линий передачи. Стоит отметить, что многочисленные вирусы: JCSMV.Iran, OCSV, CarRSV-RNA 2, RCNMV-РНК 2.Может, BBSV.Val25.Iran, GALV.nip-plefruit, TBSV.Statice, PNSV и PLPV.PV.0193 эволюционировали в соответствии с обоими механизмами: рекомбинацией и положительный выбор, между которыми может быть синергизм происходит-кольцо. Такой синергизм между рекомбинацией и естественным отбор мог сыграть важную роль в дарвиновском молекулярная эволюция. История эволюции Tombusviridae показал, что 79 образцов разделились на восемь четко отдельные кластеры, представляющие восемь родов Семейство Tombus-viridae.Из настоящего филогенетического исследования на можно сделать как минимум два таксономических следствия: (i) три ви-русы (NLVCV.Alaska, PCRPV.GR 57, PLPV.PV-0193) в настоящее время рассматривается ICTV как: один неназначенный Tombusviridae (PLPV.PV-0193) и два неклассифицированных Tombusviridae (NLVCV.Alaska и PCRPV.GR 57). Все из них следует отнести к роду Carmovirus ; (ii) В дополнение к вирусам, принадлежащим к роду Carmovirus , которые сформировали две отдельные подгруппы, члены поколения Necrovirus и Dianthovirus развивались отдельно и разделены на две отдельные подгруппы, как показано на рисунке 1.В напротив, представители родов Aureusvirus и Tombusvirus образуют отдельно единый ансамбль. Эволюционный отношения между вирусами - надежный подход к классификация. Как указано Stuart et al. (2004) (кто сообщил аналогичные результаты относительно генетической дивергенции компонентов рода Necrovirus ), сравнение полных геномов - это более сбалансированный подход, который должен обеспечить более точная схема родства.С другой стороны, это Следует отметить, что у рода Dianthovirus генетическая дивергенция между РНК 1 и 2 коррелирует с синтезированные конечные продукты и их использование вирусом для выживать. Например: молчание РНК - это управляемая малой РНК Последовательно-специфический механизм активации гена в эукариот, которые участвуют в различных биологических явления ( например, развитие , образование гетерохроматина и защита от молекулярных паразитов, таких как вирусы). Много вирусы экспрессируют супрессоры, чтобы противодействовать Противовирусная защита, опосредованная подавлением РНК.Эти молчание РНК супрессоры были идентифицированы в следующих родах: Aureusvirus , Carmovirus , Tombusvirus, и Dianthovirus (Voinnet и др. ., 1999; Qu и др. , 2003; Mérai и др. , 2005; Takeda et al. , 2005). Dianthovirus использует уникальную стратегию для подавить молчание РНК. Подавитель диантовирусов состоит из нескольких компонентов, включая P27, P88 (закодированные двумя ORF в РНК 1) и вирусной РНК (Takeda et al., г. 2005). Кроме того, изменчивость последовательности шерсти ген, кодирующий белок (РНК 1), может быть связан с взаимодействием (13)между этим структурным белком и хозяином и вектором которые сами по себе демонстрируют большое разнообразие среди диато-вирусы. Напротив, ORF в РНК 2 кодирует белок движения. Все эти факторы могут повлиять на расхождение между двумя РНК. Наконец, насколько автору известно, это крупнейшее исследование в литературе по рекомбинации потенциально присутствующие во всем геноме всех известных в настоящее время члены семейства Tombusviridae, а также положительные селекция, действующая на экспрессию белков и их филогенетическая реконструкция.Кроме того, реклассификация на основе на их предсказанную эволюционную историю. Список литературыAaziz R и Tepfer M (1999) Рекомбинация в РНК-вирусах и у устойчивых к вирусам трансгенных растений. Дж. Ген Вирол 80: 1339-1346. Alejska M, Kurzyniska-Kokorniak A, Broda M, Kierzek R and Figlerowicz M (2001) Как РНК-вирусы обмениваются своими генетический материал. Acta Bioch Polon 48: 391-407. Барот М., Орлич М., Тиль Х. Дж. И Бехер П. (2000) клеточные кодирующие последовательности NEDD8 в пестивирусе.Вирусология 278: 456-466. Бони М.Ф., Посада Д. и Фельдман М.В. (2007). непараметрический метод вывода мозаичной структуры в тройки последовательностей. Генетика 176: 1035-1047. Boulila M (2009) Структура рекомбинации и генетика родство членов семейства Bromoviridae на основании их анализ последовательности РНК 1 и 2. Гены вирусов 38: 435-444. Boulila M (2010) Предполагаемые события рекомбинации и эволюция история пяти экономически важных вирусов плодовых деревьев на основе анализа последовательности гена, кодирующего белок оболочки.Biochem Genet 48: 357-375. Fauquet CM, Mayo MA, Maniloff J, Desselberger U и Ball LA (2005) Таксономия вирусов: Классификация и номенклатура вирусы. В: Восьмой доклад Международного комитета по Таксономия вирусов. Elsevier / Academic Press, Лондон, 1259 с. Fitch WN (1971) К определению хода эволюции минимальное изменение для конкретной топологии дерева. Syst Zool 20: 406-416. Гиббс MJ, Армстронг JS и Гиббс AJ (2000) Сестринское сканирование: A Процедура Монте-Карло для оценки сигналов в рекомбинантных последовательности.Биоинформатика 16: 573-582. Грин AE и Allison RF (1994) Рекомбинация между вирусными Транскрипты РНК и трансгенных растений. Наука 263: 1423-1425. Хабили Н. и Саймонс Р. Х. (1989) Эволюционные отношения между лютеовирусами и другими РНК-вирусами растений на основе последовательности мотивов в их предполагаемой РНК-полимеразе и нуклеиновой кислоте. кислотные геликазы. Nucleic Acids Res 17: 9543-9555. Hartigan JA (1973) Минимальная мутация подходит для данного дерева. Биометрия 29: 53-63. Хасегава М., Кишино Х. и Яно Т. (1985) Свидание человека-обезьяны расщепление по молекулярным часам митохондриальной ДНК. Дж Мол Evol 22: 160-174. Джукс Т. и Кантор С. (1969) Эволюция белковых молекул. В: Munro HN (ed) Метаболизм белков млекопитающих. Академический Press, New York, pp 21-132. Кумар С., Ней М., Дадли Дж. И Тамура К. (2008) МЕГА: А биологически ориентированное программное обеспечение для эволюционного анализа ДНК и белковые последовательности. Краткая биография 9: 299-306. Lai MMC (1992) Рекомбинация РНК в вирусах животных и растений.Microbiol Rev 56: 61-79. Ларкин М.А., Blackshileds G, Brown NP, Chenna R, McGettigan PA, McWilliam H, Valentin F, Wallace IM, Wilm A, Lopez R et al. (2007) Clustal W и Clustal X v. 2.0. Биоинформатика 23: 2947-2948. Мартин Д. и Рыбицки Э. (2000) RDP: обнаружение рекомбинации среди выровненных последовательностей. Биоинформатика 16: 562-563. Мартин Д.П., Посада Д., Крэндалл К.А. и Уильямсон К. (2005a) А. модифицированный алгоритм bootscan для автоматической идентификации последовательности рекомбинации и контрольные точки рекомбинации.СПИД Res H Retrovir 21: 98-102. Мартин Д.П., Уильямсон С. и Посада Д. (2005b) RDP2: Обнаружение рекомбинации и анализ на основе выравнивания последовательностей. Биоинформатика 21: 260-262. Майдт Дж. И Ленгауэр Т. (2006) Рекко: анализ рекомбинации с помощью оптимизации затрат. Биоинформатика 22: 1064-1071. Мераи З., Кереньи З., Мольнар А., Барта Э., Валоци А., Бистрей Дж., Havelda Z, Burgyan J и Silhavy D (2005) Aureuvirus P14 является эффективным супрессором подавления РНК, который связывает двухцепочечные РНК без размерной специфичности.J Virol 79: 7217-7226. Нагай М., Сакода Й, Мори М., Хаяси М., Кида Х и Акаши Х (2003) Вставка клеточной последовательности и РНК рекомбинация в кодирующей структурный белок области цитопатогенный вирус вирусной диареи крупного рогатого скота. Дж. Ген Вирол 84Пт2: 447-452. Nei M, Gojobori T (1986) Простые методы оценки числа синонимичных и несинонимичных нуклеотидов замены. Mol Biol Evol 3: 418-426. Padidam M, Sawyer S и Fauquet CM (1999) Возможно появление новых геминивирусов путем частой рекомбинации.Вирология 265: 218-225. Посада Д. и Крэндалл К. (2001) Оценка методов для обнаружение рекомбинации по последовательностям ДНК. Компьютер моделирование. Proc Natl Acad Sci USA 98: 13757-13762. Ку Ф, Рен Т. и Моррис Т. Дж. (2003) Белок оболочки репы crin-kle virus подавляет посттранскрипционное молчание генов на шаг раннего инициирования. Дж. Вирол 77: 511-522. Riviere CJ и Rochon DM (1990) Нуклеотидная последовательность и геномная организация вируса некротической пятнистости дыни.J Gen Вирол 71: 1887-1896. Смит Дж. М. (1992) Анализ мозаичной структуры генов. Дж Мол Evol 34: 126-129. Стюарт Дж., Моффет К. и Бозарт Р.Ф. (2004) Целый геном взгляд на филогению семейства вирусов растений Tom-busviridae. Arch Virol 149: 1595-1610. Suzuki Y (2006) Статистические свойства методов обнаружения положительно выбранные аминокислотные сайты. Gene 365: 125-129. Suzuki Y и Gojobori T (1999) Метод обнаружения положительного отбор по отдельным аминокислотным сайтам . Мол Биол Эвол 16: 1315-1328.16. Таджима Ф (1989) Статистический метод проверки нейтральных Гипотеза мутации по полиморфизму ДНК. Генетика 123: 585-595. (14)Такеда А., Цукуда М., Мизумото Х., Окамото К., Кайдо М., Мисе K и Okuno T (2005) Растительный РНК-вирус подавляет РНК. подавление репликации вирусной РНК EMBO J 24: 3147-3157. Тамура К. и Ней М. (1993) Оценка количества нуклеотидные замены в контрольной области митохондриальной ДНК человека и шимпанзе.Мол Биол Эвол 10: 512-526. Тамура К., Петерсон Д., Петерсон Н., Стечер Г., Ней М. и Кумар S (2011) MEGA5: Молекулярно-эволюционная генетика Анализ с использованием максимального правдоподобия, эволюционного расстояния и методы максимальной экономии. Молекулярная биология и Evo-lution. Doi 10.1093 / molbev / msr121. Воиннет О., Пинто Ю.М. и Баулкомб округ Колумбия (1999) Подавление подавление генов: общая стратегия, используемая различными ДНК и РНК-вирусы растений. Proc Natl Acad Sci USA 96: 14147-14152. White KA и Nagy PD (2004) Достижения в области молекулярной биология tombusvirus: экспрессия генов, репликация генома и рекомбинация.Прог Нуклеиновая Кислота Res Mol Biol 78: 187-226. Worobey M и Holmes EC (1999) Эволюционные аспекты повторная комбинация в РНК-вирусах. J Gen Virol 80: 2535-2543. Заместитель редактора: Карлос Ф.М. Менк Информация о лицензии: это статья в открытом доступе, распространяемая в соответствии с условиями Лицензия Creative Commons Attribution License, которая разрешает неограниченное использование, распространение и воспроизведение на любом носителе при условии правильного цитирования оригинала. . |