


Комплексное сопротивление — это… Что такое Комплексное сопротивление?
- Комплексное сопротивление
Реактивное сопротивление — электрическое сопротивление, обусловленное передачей энергии электрическому или магнитному полю (и обратно).
Реактивное сопротивление определяет мнимую часть импеданса:
- Z = R + iX, где Z — импеданс, R — величина активного сопротивления, X — величина реактивного сопротивления, i — мнимая единица.
В зависимости от величины X какого-либо элемента электрической цепи, говорят о трёх случаях:
Величина реактивного сопротивления может быть выражена через величины индуктивного и ёмкостного сопротивлений:
Индуктивное сопротивление (XL) обусловлено возникновением ЭДС самоиндукции. Электрический ток создает магнитное поле.

Ёмкостное сопротивление (XC). Величина ёмкостного сопротивления зависит от ёмкости элемента С и также частоты протекающего тока:См. также
Активное сопротивлние

Wikimedia Foundation. 2010.
- Комплексное соединение
- Комплексные вещества
Смотреть что такое «Комплексное сопротивление» в других словарях:
комплексное сопротивление — Комплексная величина, равная отношению комплексного действующего значения синусоидального электрического напряжения на выводах пассивной электрической цепи или ее элемента к комплексному действующему значению синусоидального электрического тока в … Справочник технического переводчика
КОМПЛЕКСНОЕ СОПРОТИВЛЕНИЕ — то же, что импеданс.
Физическая энциклопедия. В 5 ти томах. М.: Советская энциклопедия. Главный редактор А. М. Прохоров. 1988 … Физическая энциклопедиякомплексное сопротивление обмотки — Импеданс измерительной обмотки, сочлененной с проводящим контролируемым изделием. [Система неразрушающего контроля. Виды (методы) и технология неразрушающего контроля. Термины и определения (справочное пособие). Москва 2003 г.] Тематики виды… … Справочник технического переводчика
комплексное сопротивление электрической цепи — Комплексная величина, равная отношению комплексного напряжения на зажимах данной цепи к комплексному току в этой цепи … Политехнический терминологический толковый словарь
Сопротивление — Сопротивление: В Викисловаре есть статья «сопротивление» Электрическое сопротивление физическая величина, характеризующая свойства проводника препятствовать прохождению электрического тока.
Сопротивление разговорное название резистора … Википедиякомплексное полное сопротивление — — [Я.Н.Лугинский, М.С.Фези Жилинская, Ю.С.Кабиров. Англо русский словарь по электротехнике и электроэнергетике, Москва, 1999 г.] Тематики электротехника, основные понятия EN vector impedance … Справочник технического переводчика
сопротивление — 3.93 сопротивление (resistance): Способность конструкции или части конструкции противостоять действию нагрузок. Источник: ГОСТ Р 54382 2011: Нефтяная и газовая промышленность. Подводные трубопроводные системы. Общие технические требования … Словарь-справочник терминов нормативно-технической документации
комплексное (электрическое) сопротивление — 154 комплексное (электрическое) сопротивление Комплексная величина, равная отношению комплексного действующего значения синусоидального электрического напряжения на выводах пассивной электрической цепи или ее элемента к комплексному действующему… … Словарь-справочник терминов нормативно-технической документации
сопротивление короткого замыкания четырехполюсника — 199 сопротивление короткого замыкания четырехполюсника Комплексное или операторное сопротивление пассивного четырехполюсника со стороны одной пары выводов, когда другая пара замкнута накоротко Источник: ГОСТ Р 52002 2003: Электротехника.
Термины… … Словарь-справочник терминов нормативно-технической документации
 Физическая энциклопедия. В 5 ти томах. М.: Советская энциклопедия. Главный редактор А. М. Прохоров. 1988 … Физическая энциклопедия
Физическая энциклопедия. В 5 ти томах. М.: Советская энциклопедия. Главный редактор А. М. Прохоров. 1988 … Физическая энциклопедия Сопротивление разговорное название резистора … Википедия
Сопротивление разговорное название резистора … Википедия Термины… … Словарь-справочник терминов нормативно-технической документации
Термины… … Словарь-справочник терминов нормативно-технической документацииКомплексное сопротивление — Большая Энциклопедия Нефти и Газа, статья, страница 1
Комплексное сопротивление
Cтраница 1
Комплексное сопротивление — полное сопротивление цепи, обладающей активным и реактивным сопротивлением, выраженное в виде комплексного числа, модуль которого равен полному сопротивлению, а аргумент равен углу сдвига фаз между током и напряжением цепи. [1]
Комплексное сопротивление или проводимость какого-либо элемента определяют его реакцию на напряжение или ток. [3]
Комплексное сопротивление может быть получено посредством последовательного го-единшия, а комплексная проводимость — посредством параллельного соединения; поэтому приведенные ф-лы служат для пересчета параллельного включения в последовательное и наоборот.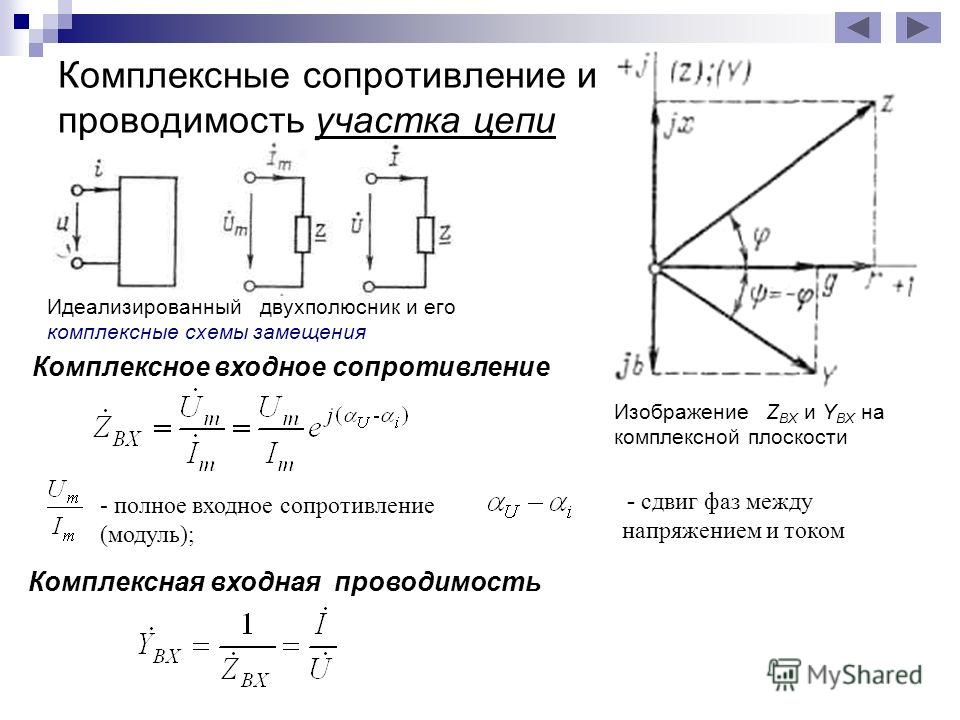
Комплексное сопротивление — полное сопротивление ( см.) цепи, которая обладает как активным, так и реактивным сопротивлением. [6]
Комплексное сопротивление является одной из важных характеристик участка ( пасивного элемента) электрической цепи переменного тока. Следует отметить, что понятие комплексного сопротивления имеет смысл лишь при условии, что в электрической цепи действуют синусоидальные напряжения и токи. Поэтому измерения комплексных сопротивлений и их составляющих должны проводиться с соблюдением этих условий. При несинусоидальных напряжении и токе результат измерения может быть получен лишь для определенных гармоник путем использования специальных избирательных устройств. [8]
Комплексное сопротивление равно 3 / 5 ом. [9]
Комплексное сопротивление равно 3 / 5 Ом.
Комплексное сопротивление Z12 является взаимным сопротивлением для входного и выходного контуров, поскольку по нему протекают одновременно входной и выходной токи. [11]
Комплексное сопротивление и его вещественная и мнимая составляющие могут быть представлены на комплексной плоскости ( рис. 12 — 6) в виде треугольника сопротивлений. [12]
Комплексное сопротивление состоит из активной и реактивной составляющих. При этом очень полезно воспользоваться аналогиями из теории электрических цепей. [13]
Комплексные сопротивления
 Эти полученные экспериментально результаты можно подтвердить строгими теоретическими выводами, что, однако, вышло бы за рамки настоящей работы.
[14]
Эти полученные экспериментально результаты можно подтвердить строгими теоретическими выводами, что, однако, вышло бы за рамки настоящей работы.
[14]Страницы: 1 2 3 4 5
Сопротивления | Цепи переменного тока
Введем теперь ряд величин, характеризующих цепь синусоидального тока.
Отношение комплексного напряжения к комплексному току называется комплексным сопротивлением:
где — отношение действующего или амплитудного напряжения соответственно к действующему или амплитудному току называется полным сопротивлением. Полное сопротивление равно модулю комплексного сопротивления. Аргумент комплексного сопротивления равен разности фаз напряжения и тока, т. е.
Комплексное сопротивление можно представить в виде
где — действительная часть комплексного сопротивления, называется активным сопротивлением; — значение мнимой части комплексного сопротивления, называется реактивным сопротивлением.
Очевидно, что
Из ( 3.23а) следует, что для последовательного контура (см. рис. 3.8) комплексное сопротивление
причем реактивное сопротивление
где
называются соответственно индуктивным и емкостным сопротивлениями.
Из ( 3.15) и ( 3.19) видно, что индуктивное сопротивление связывает между собой амплитуды или действующие значения напряжения на индуктивности и тока:
Индуктивное сопротивление прямо пропорционально частоте тока. Это объясняется тем, что напряжение на индуктивном элементе пропорционально скорости изменения тока:
Емкостное сопротивление, как следует из ( 3.16) и ( 3.20), связывает между собой амплитуды или действующие значения напряжения на емкости и тока:
Емкостное сопротивление обратно пропорционально частоте тока. Эту зависимость от частоты легко пояснить, если считать заданным напряжение на емкостном элементе, а искомой величиной ток: . Ток прямо пропорционален скорости изменения напряжения на емкостном элементе, и, следовательно, емкостное сопротивление обратно пропорционально частоте напряжения.
Напряжения на последовательно соединенных индуктивности и емкости противоположны по фазе; поэтому в (3.27) для реактивного сопротивления х сопротивления входят с различными знаками. Напряжения на индуктивности и на емкости сдвинуты по фазе относительно напряжения на сопротивлении соответственно на π/2 и —π/2. Поэтому эти сопротивления входят в Z как .
Следует обратить внимание на то, что индуктивное и емкостное сопротивления являются величинами арифметическими — положительными, а реактивное сопротивление — величина алгебраическая и может быть как больше, так и меньше нуля.
Для ветви, содержащей только индуктивность, реактивное сопротивление х равно индуктивному сопротивлению , а реактивное сопротивление х ветви, содержащей только емкость, равно емкостному сопротивлению, взятому со знаком минус, т. е. .
Заметим также, что для ветвей, каждая из которых содержит только сопротивление r, только индуктивность L или только емкость С, комплексные сопротивления соответственно равны:
Если ветвь содержит несколько последовательно соединенных резистивных, индуктивных и емкостных элементов, то при вычислении сопротивления и тока их можно заменить тремя элементами:
Элементы цепи синусоидального тока.
 Векторные диаграммы и комплексные соотношения для них. (Лекция N 4)
Векторные диаграммы и комплексные соотношения для них. (Лекция N 4)1. Резистор
Идеальный резистивный элемент не обладает ни индуктивностью, ни емкостью. Если к нему приложить синусоидальное напряжение (см. рис. 1), то ток i через него будет равен
| . | (1) |
Соотношение (1) показывает, что ток имеет ту же начальную фазу, что и напряжение. Таким образом, если на входе двухлучевого осциллографа подать сигналы u и i, то соответствующие им синусоиды на его экране будут проходить (см. рис. 2) через нуль одновременно, т.е. на резисторе напряжение и ток совпадают по фазе.
Из (1) вытекает:
;
.
Переходя от синусоидальных функций напряжения и тока к соответствующим им комплексам:
;
,
— разделим первый из них на второй:
или
. |
(2) |
Полученный результат показывает, что отношение двух комплексов есть вещественная константа. Следовательно, соответствующие им векторы напряжения и тока (см. рис. 3) совпадают по направлению.
2. Конденсатор
Идеальный емкостный элемент не обладает ни активным сопротивлением (проводимостью), ни индуктивностью. Если к нему приложить синусоидальное напряжение (см. рис. 4), то ток i через него будет равен
| . | (3) |
Полученный результат показывает, что напряжение
на конденсаторе отстает по фазе от тока на /2.
Таким образом, если на входы двухлучевого осциллографа подать сигналы
u
и i, то на его экране будет иметь
место картинка, соответствующая рис. 5.
Из (3) вытекает:
;
.
Введенный параметр называют реактивным емкостным сопротивлением конденсатора. Как и резистивное сопротивление, имеет размерность Ом. Однако в отличие от R данный параметр является функцией частоты, что иллюстрирует рис. 6. Из рис. 6 вытекает, что при конденсатор представляет разрыв для тока, а при .
Переходя от синусоидальных функций напряжения и тока к соответствующим им комплексам:
;
,
— разделим первый из них на второй:
или
| . | (4) |
В последнем соотношении — комплексное сопротивление
конденсатора. Умножение на соответствует повороту
вектора на угол по часовой стрелке. Следовательно,
уравнению (4) соответствует векторная диаграмма, представленная на рис. 7.
Умножение на соответствует повороту
вектора на угол по часовой стрелке. Следовательно,
уравнению (4) соответствует векторная диаграмма, представленная на рис. 7.
3. Катушка индуктивности
Идеальный индуктивный элемент не обладает ни активным сопротивлением, ни емкостью. Пусть протекающий через него ток (см. рис. 8) определяется выражением . Тогда для напряжения на зажимах катушки индуктивности можно записать
| . | (5) |
Полученный результат показывает, что напряжение на катушке индуктивности
опережает по фазе ток на /2.
Таким образом, если на входы двухлучевого осциллографа подать сигналы u и i, то на его экране (идеальный
индуктивный элемент) будет иметь место картинка, соответствующая рис. 9.
9.
Из (5) вытекает:
.
Введенный параметр называют реактивным индуктивным сопротивлением катушки; его размерность – Ом. Как и у емкостного элемента этот параметр является функцией частоты. Однако в данном случае эта зависимость имеет линейный характер, что иллюстрирует рис. 10. Из рис. 10 вытекает, что при катушка индуктивности не оказывает сопротивления протекающему через него току, и при .
Переходя от синусоидальных функций напряжения и тока к соответствующим комплексам:
;
,
разделим первый из них на второй:
или
| . | (6) |
В полученном соотношении — комплексное
сопротивление катушки индуктивности. Умножение на соответствует повороту вектора на угол против часовой стрелки.
Следовательно, уравнению (6) соответствует векторная диаграмма, представленная на рис. 11
Умножение на соответствует повороту вектора на угол против часовой стрелки.
Следовательно, уравнению (6) соответствует векторная диаграмма, представленная на рис. 11
4. Последовательное соединение резистивного и индуктивного элементов
Пусть в ветви на рис. 12 . Тогда
где
, причем пределы изменения .
Уравнению (7) можно поставить в соответствие соотношение
,
которому, в свою очередь, соответствует векторная диаграмма на рис. 13. Векторы
на рис. 13 образуют фигуру, называемую треугольником напряжений. Аналогично
выражение
графически может быть представлено треугольником сопротивлений (см. рис. 14), который подобен треугольнику напряжений.
5. Последовательное соединение резистивного и емкостного элементов
Последовательное соединение резистивного и емкостного элементов
Опуская промежуточные выкладки, с использованием соотношений (2) и (4) для ветви на рис. 15 можно записать
| ., | (8) |
где
, причем пределы изменения .
На основании уравнения (7) могут быть построены треугольники напряжений (см.
рис. 16) и сопротивлений (см. рис. 17), которые являются подобными.
6. Параллельное соединение резистивного и емкостного элементов
Для цепи на рис. 18 имеют место соотношения:
;
, где [См] – активная проводимость;
, где [См] – реактивная проводимость конденсатора.
Векторная диаграмма токов для данной цепи, называемая треугольником токов,
приведена на рис. 19. Ей соответствует уравнение в комплексной форме
19. Ей соответствует уравнение в комплексной форме
,
где ;
— комплексная проводимость;
.
Треугольник проводимостей, подобный треугольнику токов, приведен на рис. 20.
Для комплексного сопротивления цепи на рис. 18 можно записать
.
Необходимо отметить, что полученный результат аналогичен известному из курса физики выражению для эквивалентного сопротивления двух параллельно соединенных резисторов.
7. Параллельное соединение резистивного и индуктивного элементов
Для цепи на рис. 21 можно записать
;
, где [См] – активная проводимость;
, где [См] – реактивная проводимость катушки индуктивности.
Векторной диаграмме токов (рис. 22) для данной цепи соответствует уравнение в комплексной форме
,
где ;
— комплексная проводимость;
.
Треугольник проводимостей, подобный треугольнику токов, приведен на рис. 23.
Выражение комплексного сопротивления цепи на рис. 21 имеет вид:
.
Литература
1. Основы теории цепей: Учеб. для вузов /Г.В.Зевеке, П.А.Ионкин, А.В.Нетушил, С.В.Страхов. –5-е изд., перераб. –М.: Энергоатомиздат, 1989. -528с.
2. Бессонов Л.А. Теоретические основы электротехники: Электрические цепи. Учеб. для студентов электротехнических, энергетических и приборостроительных специальностей вузов. –7-е изд., перераб. и доп. –М.: Высш. шк., 1978. –528с.
Контрольные вопросы и задачи
1. В чем сущность реактивных сопротивлений?
2. Какой из элементов: резистор, катушку индуктивности или конденсатор – можно использовать в качестве шунта для наблюдения за формой тока?
3. Почему катушки индуктивности и конденсаторы не используются в цепях постоянного
тока?
Почему катушки индуктивности и конденсаторы не используются в цепях постоянного
тока?
4.
В ветви на рис. 12 . Определить комплексное сопротивление
ветви, если частота тока .
Ответ: .
5.
В ветви на рис. 15 . Определить комплексное сопротивление
ветви, если частота тока .
Ответ: .
6.
В цепи на рис. 18 . Определить комплексные проводимость
и сопротивление цепи для .
Ответ: ; .
7. Протекающий
через катушку индуктивности ток изменяется по закону А. Определить комплекс
действующего значения напряжения на катушке.
Ответ: .
Комплексное сопротивление и проводимость. Законы Ома
И Кирхгофа в комплексной форме. Векторные и потенциальные
(топографические) диаграммы
Рассмотрим участок цепи при последовательном соединении активного сопротивления, индуктивности и конденсатора (рис. 2.7). В схеме протекает синусоидальный ток .
2.7). В схеме протекает синусоидальный ток .
Рис. 2.7
Определим напряжение на входе схемы. В соответствии со вторым законом Кирхгофа (2.7) уравнение для мгновенных значений напряжений:
; (2.13)
. (2.14)
Подставим выражения (2.14) в уравнение (2.13). Получим:
(2.15)
Из выражения (2.15) следует, что напряжение в активном сопротивлении совпадает по фазе с током, напряжение на индуктивности опережает по фазе ток на 90o, напряжение по емкости отстает по фазе от тока на 90o.
Запишем уравнение (2.15) в комплексной форме:
(2.16)
Получим уравнение для комплексов действующих значений токов и напряжений, разделив члены (2.16) на :
. (2.17)
В уравнении (2.17) отношение комплексного напряжения к комплексному току называют комплексным сопротивлением и обозначают Z:
. (2.18)
Точку над Z не ставят, так как ее принято ставить только над комплексными величинами, отображающими синусоидальные функции времени. Как всякое комплексное число Z (2.18) может быть представлено в показательной или алгебраической форме:
Как всякое комплексное число Z (2.18) может быть представлено в показательной или алгебраической форме:
(2.19)
где z- модуль комплексного сопротивления или полное сопротивление цепи;
φ- аргумент комплексного сопротивления, — соответственно активное и реактивное сопротивление.
Для рассматриваемой схемы (рис.2.9) данные величины равны:
φ=arctg , ,
(2.20)
Знак величины реактивного сопротивления X и аргумента (фазы) зависит от соотношения индуктивного и емкостного сопротивлений.
При построении векторных диаграмм цепи возможны три режима.
1. Индуктивное сопротивление больше емкостного, величина реактивного сопротивления X и аргумента φ положительны. Цепь носит индуктивный характер. Векторы напряжений на индуктивности и емкости направлены в противоположные стороны, частично компенсируют друг друга. Вектор тока отстает от вектора напряжения на входе схемы (рис.2.8).
2. Индуктивное сопротивление меньше емкостного, величина реактивного сопротивления X и аргумента φ отрицательны. Цепь носит емкостной характер. Вектор тока опережает вектор напряжения на входе схемы (рис.2.9).
Индуктивное сопротивление меньше емкостного, величина реактивного сопротивления X и аргумента φ отрицательны. Цепь носит емкостной характер. Вектор тока опережает вектор напряжения на входе схемы (рис.2.9).
3. Индуктивное и емкостное сопротивления одинаковы. Напряжения на индуктивности и емкости полностью компенсируют друг друга. Ток в цепи совпадает по фазе с входным напряжением. В электрической цепи наступает режим резонанса напряжения (рис.2.10), который рассматривается в п.2.6.
Рис. 2.8 Рис. 2.9 Рис. 2.10
В ветвях цепи при параллельном соединении активного сопротивления, индуктивности и конденсатора (рис.2.11) удобнее выполнять расчеты с использованием комплексной проводимости.
Рис. 2.11
Отношение комплексного тока к комплексному напряжению, т.е. величину обратную комплексному сопротивлению, называют комплексной проводимостью:
(2. 21)
21)
где g, b, y — активная, реактивная и полная проводимости цепи.
. (2.22)
Единица комплексной проводимости – См (Ом -1).
Определим ток на входе схемы (рис. 2.11), считая, что на входе подключено синусоидальное напряжение .
В соответствии с первым законом Кирхгофа для мгновенных значений токов:
, (2.23)
, . (2.24)
Подставим выражения (2.24) в уравнение (2.23):
(2.25)
Из уравнения (2.25) следует, что ток в ветви с индуктивностью отстает по фазе от напряжения на 90o, ток в ветви с активным сопротивлением совпадает по фазе с напряжением, ток в ветви с емкостью опережает по фазе напряжение на 90o.
Запишем уравнение (2.25) в комплексной форме для действующих значений токов и напряжений с учетом значений проводимостей (2.22):
. (2.26)
Знак величины реактивной проводимости b и аргумента (фазы) φ зависит от величины реактивного сопротивления X. Если X положительно, то b также положительно, если X отрицательно — b отрицательно.
Если X положительно, то b также положительно, если X отрицательно — b отрицательно.
При построении векторных диаграмм цепи возможны три режима.
1. Реактивная проводимость отрицательна, величина реактивного сопротивления X и аргумента φ также отрицательны. Цепь носит емкостный характер. Вектора токов через индуктивность и емкость направлены в противоположные стороны, частично компенсируя друг друга. Вектор тока опережает вектор напряжения на входе схемы (рис.2.12).
2. Реактивная проводимость положительна, величина реактивного сопротивления X и аргумента φ также положительны. Цепь носит индуктивный характер. Вектор тока отстает от вектора напряжения на входе схемы (рис.2.13).
3. Реактивная проводимость равна нулю. Вектора токов через индуктивность и емкость направлены в противоположные стороны, компенсируя друг друга. Ток в цепи совпадает по фазе с входным напряжением. В электрической цепи наступает режим резонанса тока (рис. 2.14).
2.14).
Рис. 2.12 Рис. 2.13 Рис. 2.14
Из (2.20) следует, что модуль комплексного сопротивления можно представить как гипотенузу прямоугольного треугольника, один катет которого равен активному сопротивлению R, другой реактивному сопротивлению X (рис.2.15). Аналогично из (2.22)модуль комплексной проводимости является гипотенузой прямоугольного треугольника, один катет которого равен активной проводимости g, другой реактивной проводимости b (рис.2.16).
Рис. 2.15 Рис. 2.16
Графическая интерпретация связи между модулем полного сопротивления, активным и реактивным сопротивлениями цепи в виде прямоугольного треугольника сопротивлений аналогична графической интерпретации связи между модулем полной проводимости и ее активной и реактивной составляющими.
Комплексные уравнения (2.20) и (2. 28) представляет собой выражения закона Ома в комплексной форме:
28) представляет собой выражения закона Ома в комплексной форме:
.
Законы Кирхгофа в комплексной форме для действующих значений токов и напряжений формулируются соответственно уравнениям для мгновенных значений (2.3), (2.4).
Алгебраическая сумма комплексных значений токов в узле равна нулю (первый закон Кирхгофа)
(2.27)
Алгебраическая сумма комплексных значений ЭДС всех источников напряжения в замкнутом контуре равна алгебраической сумме комплексных напряжений на элементах этого контура (второй закон Кирхгофа)
(2.28)
Законы Кирхгофа позволяют решить две важных задачи анализа электрических цепей. Во-первых, их непосредственное применение при записи системы комплексных уравнений цепи в матричной форме и использовании математического пакета Mathcad дает возможность рассчитать без промежуточных преобразований комплексы токов ветвей. Несмотря на избыточность системы уравнений, составленной по законам Кирхгофа, этим методом возможно решение достаточно сложных цепей. Во-вторых, законы Кирхгофа являются средством проверки правильности решения любой цепи и основой для машинного построения векторных и потенциальных (векторно-топографических) диаграмм.
Во-вторых, законы Кирхгофа являются средством проверки правильности решения любой цепи и основой для машинного построения векторных и потенциальных (векторно-топографических) диаграмм.
В соответствии с первым законом Кирхгофа алгебраическая сумма комплексных значений токов в любом узле на комплексной плоскости должна иметь вид замкнутого многоугольника векторов токов или многолучевую звезду, сумма векторов которых равна нулю.
В соответствии со вторым законом Кирхгофа алгебраическая сумма комплексов падений напряжения и ЭДС на комплексной плоскости также должна иметь представление в виде замкнутого многоугольника векторов.
Для анализа и расчета разветвленных электрических цепей постоянного тока ранее рассмотрен ряд методов, позволяющих уменьшить порядок системы алгебраических уравнений по сравнению с методом непосредственного использования законов Кирхгофа. К ним относятся методы контурных токов, узловых потенциалов, двух узлов, эквивалентного генератора. Окончательные расчетные формулы этих методов получают в результате использования первого и второго законов Кирхгофа.
Указанные законы в комплексной форме справедливы для цепей синусоидального тока (2.27), (2.28). Поэтому в разветвленной электрической цепи, не имеющей элементов со взаимоиндуктивной связью, все расчетные уравнения для перечисленных методов (контурных токов, узловых потенциалов, двух узлов, эквивалентного генератора) справедливы для цепей синусоидального тока при условиях замены постоянных ЭДС, напряжений и токов (E, U, I ) комплексами их действующих значений ( , ), проводимости и сопротивления (g, R) – комплексными проводимостями и сопротивлениями (Y, Z). При этом алгебраическая система уравнений заменяется системой комплексных уравнений.
Расчет электрических цепей синусоидального тока при наличии в них элементов со взаимоиндуктивной связью имеет ряд особенностей и рассмотрен отдельно в п.2.7.
Закон Ома в комплексной форме
Для анализа электрических цепей синусоидального тока удобнее применять закон Ома в комплексной форме. Цепи синусоидального тока – линейные цепи с установившимся режимом работы, когда после окончания в них переходных процессов, падения напряжений на участках, токи в ветвях и ЭДС источников являются синусоидальными функциями времени. В обратном случае закон в такой форме неприменим.
Цепи синусоидального тока – линейные цепи с установившимся режимом работы, когда после окончания в них переходных процессов, падения напряжений на участках, токи в ветвях и ЭДС источников являются синусоидальными функциями времени. В обратном случае закон в такой форме неприменим.
В отличие от обычной формы закона Ома, в комплексной форме напряжение, токи, сопротивления и ЭДС записываются как комплексные числа. Данное нововведение основано на том, что в цепях переменного тока существуют активные и реактивные значения напряжений, токов и сопротивлений, что требует определенных корректив.
Итак, вместо активного сопротивления R, которое используется в основном в цепях постоянного тока, запишем полное (комплексное) сопротивление цепи Z. Падение напряжения, ток и ЭДС тоже становятся комплексными величинами. При практических расчетах удобнее пользоваться действующими значениями. Запишем формулу закона Ома в комплексной форме:
где- Z – комплексное (полное) сопротивление,
- Y – комплексная (полная) проводимость.

- r – активное сопротивление,
- x – реактивное сопротивление,
- z – полное сопротивление,
- g – активная проводимость,
- b – реактивная проводимость,
- y – полная проводимость,
- j – комплексная единица, j=√(-1).
Решение задач
По заданной схеме определить полное сопротивление цепи, токи (I_1 ) ̇, (I_2 ) ̇, (I_3 ) ̇. U = 120 В, xC1 = 100 Ом, xL2 = 50 Ом, xC3 = 50 Ом, r1 = 25 Ом, r2 = 20 Ом.
| Дано: | Решение: |
|---|---|
|
|
Сопротивление конденсатора
Господа, сегодняшнюю статью можно считать в некотором роде продолжением предыдущей. Сначала я даже хотел поместить весь этот материал в одну статью. Но его получилось довольно много, на горизонте были новые проекты, и я в итоге разделил его на две. Итак, сегодня мы поговорим про сопротивление конденсатора переменному току. Мы получим выражение, по которому можно будет рассчитать, чему равно сопротивление любого конденсатора, включенного в цепь с переменным током, а в конце статьи рассмотрим несколько примеров такого расчета.
Сначала я даже хотел поместить весь этот материал в одну статью. Но его получилось довольно много, на горизонте были новые проекты, и я в итоге разделил его на две. Итак, сегодня мы поговорим про сопротивление конденсатора переменному току. Мы получим выражение, по которому можно будет рассчитать, чему равно сопротивление любого конденсатора, включенного в цепь с переменным током, а в конце статьи рассмотрим несколько примеров такого расчета.
Сразу оговорюсь про одну важную вещь. Вообще говоря, реальный конденсатор обладает помимо емкостного сопротивления еще резистивным и индуктивным. На практике все это надо обязательно учитывать, потому что возможны ситуации (обычно связанные с ростом частоты сигнала), когда конденсатор перестает быть конденсатором и превращается… в некое подобие катушки индуктивности . При проектировании схем этот момент обязательно надо иметь в виду. Согласитесь, господа, крайне неприятно поставить в схему конденсатор и потом столкнуться с тем, что из-за высокой частоты он ведет себя и не как конденсатор вовсе, а как самый настоящий дроссель. Это, безусловно, очень важная тема, но сегодня речь пойдет не о ней. В сегодняшней статье мы будем говорить непосредственно про емкостное сопротивление конденсатора. То есть мы будем считать его идеальным, без каких бы то ни было паразитных параметров вроде индуктивности или активного сопротивления.
Это, безусловно, очень важная тема, но сегодня речь пойдет не о ней. В сегодняшней статье мы будем говорить непосредственно про емкостное сопротивление конденсатора. То есть мы будем считать его идеальным, без каких бы то ни было паразитных параметров вроде индуктивности или активного сопротивления.
Давайте представим, что у нас есть конденсатор, который включен в цепь с переменным током. В цепи больше нет никаких компонентов, только один конденсатор и все (рисунок 1).
Рисунок 1 – Конденсатор в цепи переменного тока
К его обкладкам приложено некоторое переменное напряжение U(t), и через него течет некоторый ток I(t). Зная одно, можно без проблем найти другое. Для этого надо всего лишь вспомнить прошлую статью про конденсатор в цепи переменного тока, там мы про все это довольно подробно говорили. Будем полагать, что ток через конденсатор изменяется по синусоидальному закону вот так
В прошлой статье мы пришли к выводу, что если ток изменятся вот по такому закону, то напряжение на конденсаторе должно меняться следующим образом
Пока что ничего нового мы не записали, это все дословное повторение выкладок из предыдущей статьи. А сейчас самое время их немного преобразовать, придать им чуть другой облик. Если говорить конкретно, то нужно перейти к комплексному представлению сигналов! Помните, на эту тему была отдельная статья? В ней я говорил, что она нужна для понимания некоторых моментов в дальнейших статьях. Вот как раз и наступил тот момент, когда пора вспомнить все эти хитрые мнимые единицы. Если говорить конкретно, то сейчас нам потребуется показательная запись комплексного числа. Как мы помним из статьи про комплексные числа в электротехнике, если у нас есть синусоидальный сигнал вида
А сейчас самое время их немного преобразовать, придать им чуть другой облик. Если говорить конкретно, то нужно перейти к комплексному представлению сигналов! Помните, на эту тему была отдельная статья? В ней я говорил, что она нужна для понимания некоторых моментов в дальнейших статьях. Вот как раз и наступил тот момент, когда пора вспомнить все эти хитрые мнимые единицы. Если говорить конкретно, то сейчас нам потребуется показательная запись комплексного числа. Как мы помним из статьи про комплексные числа в электротехнике, если у нас есть синусоидальный сигнал вида
то его можно представить в показательной форме вот так
Почему это так, откуда взялось, что здесь какая буковка значит – обо всем уже подробно говорили. Для повторения можно перейти по ссылке и еще раз со всем ознакомиться.
Давайте-ка теперь применим это комплексное представление для нашей формулы напряжения на конденсаторе. Получим что-то типа такого
Теперь, господа, я хотел бы вам рассказать еще про один интересный момент, который, наверное, следовало бы описать в статье про комплексные числа в электротехнике. Однако тогда я про него как-то позабыл, поэтому давайте рассмотрим его сейчас. Давайте представим, что t=0. Это приведет к исключению из расчетов времени и и частоты, и мы переходим к так называемым комплексным амплитудам сигнала. Безусловно, это не значит, что сигнал из переменного становится постоянным. Нет, он все так же продолжает изменяться по синусу с той же самой частотой. Но бывают моменты, когда частота нам не очень важна, и тогда лучше от нее избавиться и работать только с амплитудой сигнала. Сейчас как раз такой момент. Поэтому полагаем t=0 и получаем комплексную амплитуду напряжения
Однако тогда я про него как-то позабыл, поэтому давайте рассмотрим его сейчас. Давайте представим, что t=0. Это приведет к исключению из расчетов времени и и частоты, и мы переходим к так называемым комплексным амплитудам сигнала. Безусловно, это не значит, что сигнал из переменного становится постоянным. Нет, он все так же продолжает изменяться по синусу с той же самой частотой. Но бывают моменты, когда частота нам не очень важна, и тогда лучше от нее избавиться и работать только с амплитудой сигнала. Сейчас как раз такой момент. Поэтому полагаем t=0 и получаем комплексную амплитуду напряжения
Давайте раскроем скобки в экспоненте и воспользуемся правилами работы с показательными функциями.
Итак, у нас имеется три множителя. Будем разбираться со всеми по порядку. Объединим первые два и запишем выражение следующего вида
Что мы вообще такое записали? Правильно, комплексную амплитуду тока через конденсатор. Теперь выражение для комплексной амплитуды напряжения принимает вид
Теперь выражение для комплексной амплитуды напряжения принимает вид
Результат, к которому мы стремимся, уже близок, но остается еще один не очень приятный множитель с экспонентой. Как с ним быть? А, оказывается, очень просто. И снова нам на помощь придет статья по комплексным числам в электротехнике, не зря ж я ее писал . Давайте преобразуем этот множитель, воспользовавшись формулой Эйлера:
Да, вся эта хитрая экспонента с комплексными числами в показателе превращается всего лишь в мнимую единичку, перед которой стоит знак минус. Согласен, возможно, осознать это не так просто, но тем не менее математика говорит, что это так. Поэтому результирующая формула у нас принимает вид
Давайте выразим из этой формулы ток и приведем выражение к виду, соответствующему закону Ома. Получим
Как мы помним из статьи про закон Ома, у нас ток равнялся напряжению, деленному на сопротивление. Так вот, здесь практически то же самое! Ну, за исключением того, что у нас ток и напряжение – переменные и представлены через комплексные амплитуды. Кроме того, не забываем, что ток течет у нас через конденсатор. Поэтому, выражение, которое стоит в знаменателе, можно рассматривать как емкостное сопротивление конденсатора переменному току:
Кроме того, не забываем, что ток течет у нас через конденсатор. Поэтому, выражение, которое стоит в знаменателе, можно рассматривать как емкостное сопротивление конденсатора переменному току:
Да, выражение для сопротивления конденсатора имеет вот такой вот вид. Оно, как вы можете заметить, комплексное. Об этом свидетельствует буковка j в знаменателе дроби. А что значит эта комплексность? На что она влияет и что показывает? А показывает она, господа, исключительно сдвиг фаз в 90 градусов между током и напряжением на конденсаторе. А именно, ток на 90 градусов опережает напряжение. Этот вывод не является для нас новостью, про все это было подробно рассказано в прошлой статье. Чтобы это лучше осознать, надо теперь мысленно пройтись от полученной формулы вверх к тому моменту, где у нас это j возникло. В процессе подъема вы увидите, что мнимая единица j возникло из формулы Эйлера из-за того, что там был компонент . Формула Эйлера у нас возникла из комплексного представления синусоиды. А в исходной синусоиде как раз был заложен сдвиг фазы в 90 градусов тока относительно напряжения. Как-то так. Вроде все логично и ничего лишнего не возникло.
Формула Эйлера у нас возникла из комплексного представления синусоиды. А в исходной синусоиде как раз был заложен сдвиг фазы в 90 градусов тока относительно напряжения. Как-то так. Вроде все логично и ничего лишнего не возникло.
Теперь может возникнуть два совершенно логичных вопроса: как работать с таким представлением и в чем его выгода? Да и вообще, пока лишь какие-то дико абстрактные буковки и нифига не ясно, как взять и оценить сопротивление какого-нибудь конкретно конденсатора, который мы купили в магазине и воткнули в схему. Давайте разбираться постепенно.
Как мы уже говорили, буковка j в знаменателе говорит нам лишь о сдвиге фаз тока и напряжения. Но она не влияет на амплитуды тока и напряжения. Соответственно, если сдвиг фаз нас не интересует, то можно исключить эту буковку из рассмотрения и получить более простое выражение абсолютно без всяких комплексностей:
Согласитесь, жить стало чуточку легче. Это выражение позволяет рассчитать сопротивление конденсатора для конкретной емкость и частоты сигнала. Заметьте, господа, интересный факт. Сопротивление конденсатора, оказывается, зависит не только от самого конденсатора (а именно его емкости), но и от частоты протекающего тока. Если вспомнить обычные резисторы, то в них у нас сопротивление зависело только от самого резистора, материала, формы и всего такого прочего, но не зависело от частоты (разумеется, мы говорим сейчас про идеальные резисторы, без всяких паразитных параметров). Здесь все по-другому. Один и тот же конденсатор на разной частоте будет иметь разное сопротивление и через него будет течь ток разной амплитуды при одной и той же амплитуде напряжения.
Заметьте, господа, интересный факт. Сопротивление конденсатора, оказывается, зависит не только от самого конденсатора (а именно его емкости), но и от частоты протекающего тока. Если вспомнить обычные резисторы, то в них у нас сопротивление зависело только от самого резистора, материала, формы и всего такого прочего, но не зависело от частоты (разумеется, мы говорим сейчас про идеальные резисторы, без всяких паразитных параметров). Здесь все по-другому. Один и тот же конденсатор на разной частоте будет иметь разное сопротивление и через него будет течь ток разной амплитуды при одной и той же амплитуде напряжения.
Что еще мы можем сказать, глядя на эту формулу? Например, то, что чем больше частота сигнала, тем меньше для него сопротивление конденсатора. И чем больше емкость конденсатора, тем меньше его сопротивление переменному току.
По аналогии с резисторами, сопротивление конденсаторов измеряется все так же в Омах. Однако всегда следует помнить, что это немного другое сопротивление, его называют реактивным. И другое оно в первую очередь из-за того самого пресловутого j в знаменателе, то есть из-за сдвига фазы. У «обычных» (которые называют активными) Омов такого сдвига нет, там напряжение четко совпадает по фазе с током. Давайте построим график зависимости сопротивления конденсатора от частоты. Для определенности емкость конденсатора возьмем фиксированной, скажем, 1 мкФ. График представлен на рисунке 2.
И другое оно в первую очередь из-за того самого пресловутого j в знаменателе, то есть из-за сдвига фазы. У «обычных» (которые называют активными) Омов такого сдвига нет, там напряжение четко совпадает по фазе с током. Давайте построим график зависимости сопротивления конденсатора от частоты. Для определенности емкость конденсатора возьмем фиксированной, скажем, 1 мкФ. График представлен на рисунке 2.
Рисунок 2 (кликабельно) – Зависимость сопротивления конденсатора от частоты
На рисунке 2 мы видим, что сопротивление конденсатора переменному току убывает по закону гиперболы.
При стремлении частоты к нулю (то есть фактически при стремлении переменного току к постоянному) сопротивление конденсатора стремится к бесконечности. Это и логично: мы все помним, что для постоянного тока конденсатор фактически представляет собой разрыв цепи. На практике оно, конечно, не бесконечно, а ограничено сопротивлением утечки конденсатора. Тем не менее, оно все равно очень велико и часто его и считают бесконечно большим.
Тем не менее, оно все равно очень велико и часто его и считают бесконечно большим.
При стремлении частоты к бесконечности, сопротивление конденсатора стремится к нулю. Это все в теории, конечно. На практике реальный конденсатор обладает рядом паразитных параметров (в частности, паразитная индуктивности и сопротивление утечки), из-за чего сопротивление уменьшается только лишь до некоторой определенной частоты, а потом начинает наоборот расти. Но об этом более подробно в другой раз.
Есть еще один вопрос, который хотелось бы обговорить, прежде чем начинать рассмотрение примеров. Зачем вообще писать букву j в знаменателе сопротивления? Не достаточно ли просто всегда помнить про сдвиг фаз, а в записи использовать числа без этой мнимой единицы? Оказывается, нет. Представим себе цепь, где одновременно присутствуют резистор и конденсатор. Скажем, они соединены последовательно. И вот тут-то как раз мнимая единичка рядом с емкостью не позволит просто так взять и сложить активное и реактивное сопротивление в одно действительное число. Общее сопротивление такой цепочки будет комплексным, причем состоящим как из действительной части, так и из мнимой. Действительная часть будет обусловлена резистором (активными сопротивлением), а мнимая – емкостью (реактивным сопротивлением). Впрочем, это все тема для другой статьи, сейчас не будем в это углубляться. Давайте лучше перейдем к примерам.
Общее сопротивление такой цепочки будет комплексным, причем состоящим как из действительной части, так и из мнимой. Действительная часть будет обусловлена резистором (активными сопротивлением), а мнимая – емкостью (реактивным сопротивлением). Впрочем, это все тема для другой статьи, сейчас не будем в это углубляться. Давайте лучше перейдем к примерам.
Пусть у нас есть конденсатор емкостью, скажем C=1 мкФ. Требуется определить его сопротивление на частоте f1=50 Гц и на частоте f2=1 кГц. Кроме того, следует определить амплитуду тока с учетом того, что амплитуда приложенного к конденсатору напряжения равна Um=50 В. Ну и построить графики напряжения и тока.
Собственно, задачка эта элементарная. Подставляем циферки в формулу для сопротивления и получаем для частоты f1=50 Гц сопротивление, равное
А для частоты f2=1 кГц сопротивление будет
По закону Ома находим величину амплитуды тока для частоты f1=50 Гц
Аналогично для второй частоты f2=1 кГц
Теперь мы легко можем записать законы изменения тока и напряжения, а также построить графики для этих двух случаев. Полагаем, что напряжение у нас изменяется по закону синуса для первой частоты f1=50 Гц следующим образом
Полагаем, что напряжение у нас изменяется по закону синуса для первой частоты f1=50 Гц следующим образом
А для второй частоты f2=1 кГц вот так
Дальше мы помним, что ток в конденсаторе опережает напряжение на . Поэтому с учетом этого можем записать закон изменения тока через конденсаторы для первой частоты f1=50 Гц
и для частоты f2=1 кГц
Графики тока и напряжения для частоты f1=50 Гц представлены на рисунке 3
Рисунок 3 (кликабельно) – Напряжение на конденсаторе и ток через конденсаторе, f1=50 Гц
Графики тока и напряжения для частоты f2=1 кГц представлены на рисунке 4
Рисунок 4 (кликабельно) – Напряжение на конденсаторе и ток через конденсаторе, f2=1 кГц
Итак, господа, мы сегодня познакомились с таким понятием, как сопротивление конденсатора переменному току, научились его считать и закрепили полученные знания парочкой примеров.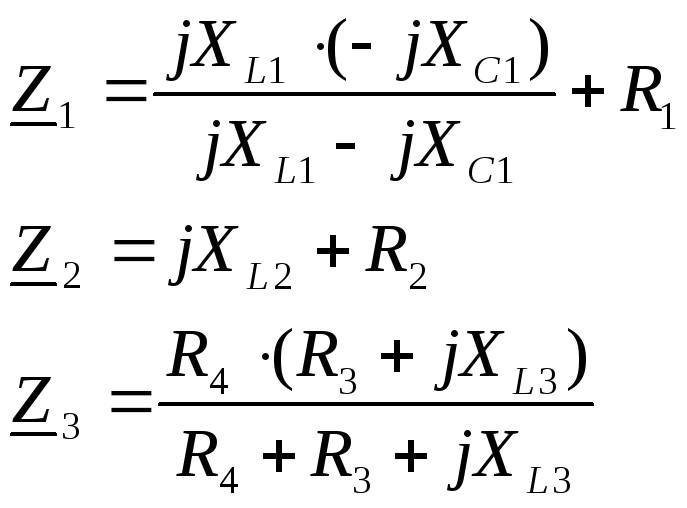 На сегодня все. Спасибо что прочитали, всем огромной удачи и пока!
На сегодня все. Спасибо что прочитали, всем огромной удачи и пока!
Вступайте в нашу группу Вконтакте
Вопросы и предложения админу: This email address is being protected from spambots. You need JavaScript enabled to view it.
границ | Применение дистанционного зондирования для фенотипирования устойчивости комплекса пятен дегтя на кукурузе
Введение
Комплекс дегтярных пятен (TSC) — это серьезное заболевание листьев кукурузы во многих регионах Латинской Америки. Заболевание вызывается взаимодействием двух грибковых патогенов: Phyllachora maydis Maubl. и Monographella maydis Müller & Samuels. Третий гриб, Coniothyrium phyllachorae Maubl. также часто ассоциируется с TSC (Hock et al., 1989). Первоначальные симптомы болезни проявляются в виде темных овальных стромов или стромы неправильной формы P. maydis , прорывающихся через эпидермис нижних и центральных листьев (рис. 1А). Приблизительно через 2 недели область, окружающая стромы, становится хлоротичной, образуя ореол, который часто называют типичным симптомом «рыбьего глаза» при TSC, который вызывается M. maydis (рис. 1B). Примерно в течение 1 месяца симптомы болезни прогрессируют с нижних листьев на верхние.В то время как P. maydis является облигатным паразитом, M. maydis считается эндофитом или факультативным паразитом, вызывающим обширный хлороз в присутствии P. maydis (Dittrich et al., 1991; Hock et al. , 1992). Неизвестно, могут ли M. maydis и P. maydis присутствовать в качестве патогенов в растении независимо, или заражение кукурузы вызвано только их одновременным совместным появлением. Хотя C. phyllachorae часто выделяют из листьев, зараженных P.maydis и M. maydis , его роль в TSC до сих пор не ясна, и считается, что он является гиперпаразитом или микопаразитом, без явного проявления симптомов хозяина (Ceballos and Deutsch, 1992; Hock et al.
1А). Приблизительно через 2 недели область, окружающая стромы, становится хлоротичной, образуя ореол, который часто называют типичным симптомом «рыбьего глаза» при TSC, который вызывается M. maydis (рис. 1B). Примерно в течение 1 месяца симптомы болезни прогрессируют с нижних листьев на верхние.В то время как P. maydis является облигатным паразитом, M. maydis считается эндофитом или факультативным паразитом, вызывающим обширный хлороз в присутствии P. maydis (Dittrich et al., 1991; Hock et al. , 1992). Неизвестно, могут ли M. maydis и P. maydis присутствовать в качестве патогенов в растении независимо, или заражение кукурузы вызвано только их одновременным совместным появлением. Хотя C. phyllachorae часто выделяют из листьев, зараженных P.maydis и M. maydis , его роль в TSC до сих пор не ясна, и считается, что он является гиперпаразитом или микопаразитом, без явного проявления симптомов хозяина (Ceballos and Deutsch, 1992; Hock et al. , 1992). В случае тяжелых эпидемий TSC хлоротичные ореолы сливаются, и все растение может стать некротизированным примерно в течение недели. Hock et al. (1989) предположили возможное участие производства фитотоксинов в TSC как причину быстрого эффекта «сжигания» листвы. Было высказано предположение, что оптимальная температура для развития болезни составляет 16–18 ° C (± 5–7 ° C) при среднемесячном количестве осадков 150 мм и 10–20 туманных дней в месяц (Hock et al., 1989).
, 1992). В случае тяжелых эпидемий TSC хлоротичные ореолы сливаются, и все растение может стать некротизированным примерно в течение недели. Hock et al. (1989) предположили возможное участие производства фитотоксинов в TSC как причину быстрого эффекта «сжигания» листвы. Было высказано предположение, что оптимальная температура для развития болезни составляет 16–18 ° C (± 5–7 ° C) при среднемесячном количестве осадков 150 мм и 10–20 туманных дней в месяц (Hock et al., 1989).
Рисунок 1 . Симптомы комплекса пятен дегтя (TSC) на растениях кукурузы: (A) стромы Phyllachora maydis первоначально появляются на листьях; (B) хлоротичные и некротические пятна, вызванные Monographella maydis , окружают стромы P. maydis ~ 2 недели спустя, вызывая так называемый симптом «рыбьего глаза».
Комплекс болезни был зарегистрирован в нескольких странах Латинской Америки и, как полагали, ограничивался тропическими районами региона (Hock et al. , 1989). Большая часть информации о потерях урожая, вызванных TSC, носит анекдотический характер. Хок (1988) упомянул потерю 30% урожая кукурузы, но не уточнил детали исследования. В другом исследовании Hock et al. (1995) сообщили о потерях урожая от 11 до 25% в Поса-Рика, Веракрус, Мексика, хотя подробности этого исследования также не были раскрыты. Bajet et al. (1994) сообщили о 46% потерях в своем исследовании эффективности фунгицидов в том же месте. Насколько нам известно, последнее исследование является единственным подробным отчетом о потерях урожая для TSC, и он не обновлялся.
, 1989). Большая часть информации о потерях урожая, вызванных TSC, носит анекдотический характер. Хок (1988) упомянул потерю 30% урожая кукурузы, но не уточнил детали исследования. В другом исследовании Hock et al. (1995) сообщили о потерях урожая от 11 до 25% в Поса-Рика, Веракрус, Мексика, хотя подробности этого исследования также не были раскрыты. Bajet et al. (1994) сообщили о 46% потерях в своем исследовании эффективности фунгицидов в том же месте. Насколько нам известно, последнее исследование является единственным подробным отчетом о потерях урожая для TSC, и он не обновлялся.
Несмотря на то, что TSC исторически встречались только в некоторых тропических районах Латинской Америки, P. maydis (отдельно, т. Е. Не в ассоциации с M. maydis ) был впервые обнаружен в нескольких местах на Среднем Западе США в 2015 г. (Bissonnette, 2015; Ruhl et al., 2016; Wise et al., 2016). Помимо повторного подтверждения в тех же местах в 2016 году, патоген также был зарегистрирован во Флориде (Bradley, 2016; Hansen et al. , 2016; Miller, 2016) и Миннесоте в 2017 и 2018 годах (Dr.Мартин Чилверс, личное сообщение). Сообщений о потерях урожая, вызванных только P. maydis , пока не поступало; однако Mottaleb et al. (2018) выдвинули гипотезу, что если бы TSC действительно образовался в США в результате ассоциации M. maydis с P. maydis , потеря всего лишь 1% общенационального производства зерна составила бы 1,5 миллиона метрических тонн. 231,6 млн долларов США.
, 2016; Miller, 2016) и Миннесоте в 2017 и 2018 годах (Dr.Мартин Чилверс, личное сообщение). Сообщений о потерях урожая, вызванных только P. maydis , пока не поступало; однако Mottaleb et al. (2018) выдвинули гипотезу, что если бы TSC действительно образовался в США в результате ассоциации M. maydis с P. maydis , потеря всего лишь 1% общенационального производства зерна составила бы 1,5 миллиона метрических тонн. 231,6 млн долларов США.
На данный момент фенотипирование устойчивости кукурузы к ТСК проводится только в полевых условиях из-за отсутствия надежной техники инокуляции в теплице.В Международном центре улучшения кукурузы и пшеницы (CIMMYT) оценка болезней проводится по шкале от 1 до 5, в которой одна очень устойчивая (нет симптомов болезни), а пять — очень восприимчивы, со всеми хлоротичными и некротическими листьями. Оценка заболевания проводится путем визуального наблюдения каждые 10–14 дней, начиная с момента цветения, обычно от трех до четырех раз, в зависимости от тяжести заболевания и скорости его развития. Однако в многочисленных предыдущих исследованиях сообщалось о возможных неточностях в оценках болезней зрения из-за предвзятости оценщиков (Nutter, 1993; Newton and Hackett, 1994; Parker et al., 1995; Стеддом и др., 2005; Bock et al., 2008, 2010). Несоответствия в оценке заболевания могут возникать в одном и том же исследовании как между разными оценщиками болезни, так и между разными оценками, выставленными одним и тем же оценщиком в данном испытании. Кроме того, визуальная оценка может занять много времени и денег, требуя высококвалифицированного и опытного технического персонала.
Однако в многочисленных предыдущих исследованиях сообщалось о возможных неточностях в оценках болезней зрения из-за предвзятости оценщиков (Nutter, 1993; Newton and Hackett, 1994; Parker et al., 1995; Стеддом и др., 2005; Bock et al., 2008, 2010). Несоответствия в оценке заболевания могут возникать в одном и том же исследовании как между разными оценщиками болезни, так и между разными оценками, выставленными одним и тем же оценщиком в данном испытании. Кроме того, визуальная оценка может занять много времени и денег, требуя высококвалифицированного и опытного технического персонала.
Изображения, полученные с помощью дистанционного зондирования (RS), сыграли важную роль в фенотипировании растений в различных средах и для различных культур.Цель технологии — минимизировать трудозатраты, сократить время, необходимое для фенотипирования, и повысить точность фенотипических данных. Поверхность листа по-разному поглощает, пропускает или отражает световое излучение в зависимости от его внутренней структуры, химического состава и стадии развития растения. Спектральные радиометры обнаруживают длины электромагнитных волн, выходящие за рамки видимых человеческим глазом, например отражательную способность в инфракрасном спектре. Эта информация может быть объединена, чтобы определить конкретные особенности растений, которые могут быть не заметны в видимом спектре.Таким образом, измерение спектральной отражательной способности можно использовать для понимания состояния здоровья растений или для количественной оценки степени заболевания пораженных частей растения (Simko et al., 2017). Однако следует также принимать во внимание сопутствующие эффекты, вызванные абиотическими стрессами. Такие факторы, как водный стресс и стресс, связанный с питательными веществами, также могут снизить фотосинтетическую активность растения, что, в свою очередь, влияет на коэффициент отражения листьев на уровне полога.
Лабораторная спектроскопия использовалась для обнаружения различных болезней ряда сельскохозяйственных культур (Bauriegel et al., 2011а, б; Mahlein et al., 2012, 2017; Bergsträsser et al., 2015; Куска и др., 2015). Кроме того, низковысотная полевая спектроскопия, включая многоспектральные и гиперспектральные удаленные изображения, использовалась для фенотипирования устойчивости к болезням у различных культур. К ним относятся мучнистая роса и листовая ржавчина пшеницы; Болезнь Хуанлунбин на апельсиновых деревьях; корневая гниль хлопка; Flavescence dorée и болезнь ствола винограда на винограднике; фитофтороз картофеля; и Xylella fastidiosa оливкового цвета (Franke and Menz, 2007; Yang et al., 2010; Гарсия-Руис и др., 2013; Khaled et al., 2017; Зарко-Техада и др., 2018; Альбетис и др., 2019; Franceschini et al., 2019). В большинстве этих исследований использовались видимые (VI, в основном области зеленого и красного спектра) и ближний инфракрасный (NIR) спектры с использованием гиперспектральных и / или мультиспектральных датчиков. Кроме того, в большинстве этих исследований использовались классификационные алгоритмы машинного обучения для отличия больных растений от здоровых. В целом точность варьировала от 50 до 90% в зависимости от стадии развития болезни.В некоторых исследованиях спектральная область с красным краем, которая обычно связана со структурными особенностями кроны и листьев (например, индекс площади листа, LAI), оказалась надежным индикатором для различения различных уровней тяжести заболевания (Garcia-Ruiz et al., 2013; Franceschini et al., 2019). Несмотря на их успешное применение для фенотипирования различных болезней растений, эти технологии RS никогда не использовались для фенотипирования болезней листьев кукурузы, включая TSC.
Первой целью этого исследования было изучить потенциал мультиспектральной и тепловизионной визуализации с использованием беспилотного летательного аппарата для определения фенотипа устойчивости кукурузы к TSC и сравнить эффективность этого метода с традиционной оценкой визуальных заболеваний.Вторая цель заключалась в оценке потенциальных потерь урожая зерна для TSC во влажной низменной тропической среде в Мексике.
Материалы и методы
Растительный материал
Двадцать пять тропических и субтропических гибридов кукурузы были отобраны для эксперимента (Таблица 1). Все гибриды были ранее оценены на агрономические характеристики и устойчивость к TSC во многих местах в Мексике (данные не показаны). Гибриды включали два устойчивых (CLTHW13007 и CLTHW13008) и два восприимчивых (DTMA-112 / DTMA-229 и DTMA-217 / DTMA-207) контроля, и их статус был основан на их реакции на заболевание во время предыдущих оценок устойчивости к TSC. СИММИТ (данные не показаны).
Таблица 1 . Потеря урожая зерна, вызванная комплексом пятен смолы кукурузы, рассчитанная как процентная разница урожайности зерна (т / га) между фунгицидными и нефунгицидными обработками в течение циклов выращивания 2016 и 2017 годов в Международном центре улучшения кукурузы и пшеницы (CIMMYT), Мексика.
Прививки и оценка болезней
Эксперимент проводился во время вегетационных циклов зима-весна 2016 и 2017 гг. На экспериментальной станции CIMMYT Agua Fria на севере штата Пуэбла, Мексика (20.45 ° с.ш., 97,64 ° з.д.) на высоте 110 м над уровнем моря (рисунок 2). Типичное годовое количество осадков на станции составляет ~ 1200 мм, а температура воздуха колеблется от 5 до 42 ° C в течение зимнего вегетационного цикла (ноябрь – апрель) при средней относительной влажности 85%. Почвы — суглинки с pH 7,5–8,5. Низкие температуры, высокая относительная влажность и продолжительное увлажнение листьев в этот период также способствуют развитию северного фитофтороза (НЛБ) кукурузы, вызываемого Setosphaeria turcica (Luttrell) Леонард и Саггс [anamorph Exserohilum turcicum (Passerini) Леонард. и предлагает].Чтобы избежать коинфекции NLB и способствовать развитию TSC, посев отложили на 2 месяца, а посевы были посажены в конце января. Это способствовало оптимальным условиям развития TSC при одновременном снижении риска инфекции NLB.
Рисунок 2 . Место проведения эксперимента в Международном центре улучшения кукурузы и пшеницы (CIMMYT), экспериментальная станция Agua Fria в штате Пуэбла, Мексика (A) . Цветное инфракрасное изображение (790, 660, 550 нм) гибридов кукурузы в экспериментальных испытаниях при обработке фунгицидами (A1) и нефунгицидами (A2) комплекса пятен смолы кукурузы.Данные изображения были извлечены из двух полигонов из двух центральных строк на каждом графике (B) .
Инокуляцию проводили следующим образом: смесь нескольких TSC-чувствительных, но устойчивых к NLB генотипов кукурузы высаживали за 30–45 дней до эксперимента на небольшом участке (~ 10–20 м. 2 ) в том же месте, что и эксперимент. Эти растения были инфицированы TSC естественным путем на поздних стадиях развития растений и предоставили начальный инокулят для испытания. Листья (всего ~ 100–150) этих восприимчивых растений с полностью выраженными симптомами TSC были собраны, погружены в воду на несколько секунд для увлажнения поверхности листьев и помещены в 200-литровый пластиковый бочонок, закрытый крышкой, и оставляют инкубировать в тени в течение ~ 24 часов.Затем бочку наполняли водой, листья слегка встряхивали и полученную суспензию спор фильтровали через грубое сито для удаления крупных остатков листьев. Твин-20 (Sigma Aldrich, Сент-Луис, Миссури, США) добавляли к суспензии вода / споры (1 мл на 15 л посевного материала) в качестве поверхностно-активного вещества. Суспензию спор распыляли на листву кукурузы с помощью ручного опрыскивателя после 18–7 часов вечера, чтобы использовать период увлажнения листьев, вызванного росой, в течение ночи. Инокуляцию проводили дважды: первый — до стадии метелки, а второй — через 7 дней.
Оценки болезней проводились три-четыре раза за вегетационный период с 10-дневными интервалами между каждой оценкой, начиная с цветения. Оценка болезней проводилась на всех делянках с использованием шкалы оценки болезней от 1 до 5, в которой: 1 = высокая устойчивость или реакция, близкая к иммунной, при почти 0% зараженных листьев и без видимых стромат P. maydis ; 2 = реакция от устойчивости к реакции средней устойчивости, при которой 1-25% площади листа поражено несколькими разбросанными стромами P.maydis ; 3 = реакция от умеренной до умеренно чувствительной с поражением 26–50% площади листа с умеренной плотностью хлоротичных поражений с хорошо развитыми симптомами «рыбий глаз», индуцированными M. maydis ; 4 = чувствительная реакция с 51–75% листьев, пораженных большими сросшимися хлоротичными и некротическими пятнами; 5 = высокочувствительная реакция с 76–100% листьев, пораженных обширным некрозом и часто преждевременным старением растений.
Экспериментальный образец
Урожай был посажен двумя соседними блоками в форме квадратной решетки, каждый с тремя рандомизированными повторностями, в конце января 2016 года и январе 2017 года.Один блок был без TSC (заболевание, контролируемое применением фунгицидов, в данном документе именуемое «обработка фунгицидами»; фиг. 2A1), а другой был TSC-инфицированным (без фунгицидного контроля, в данном документе именуемым «нефунгицидным лечением», фиг. 2A2 ). Опытные делянки состояли из четырех рядов длиной 5 м, расположенных на расстоянии 60 см друг от друга, с междурядьем между растениями ~ 20–25 см. Участки удобряли N-P-K в дозе 150-80-30 в соответствии с рекомендуемыми дозами, основанными на анализе почвы. Орошение применялось по мере необходимости.Чтобы предотвратить перенос фунгицидов между блоками, для разделения двух блоков использовались четыре ряда близко посаженных растений кукурузы (заполнители) и пустая полоса земли шириной 1,5 м. В блоке обработки фунгицидами болезнь контролировали с помощью фунгицида Priori Xtra TM (Азоксистробин 18.2 + Ципроконазол 7.27, Syngenta Crop Protection, Гринсборо, Северная Каролина) из расчета 1 л / га. Фунгицид применяли с помощью ручных опрыскивателей с 7-10-дневными интервалами, по крайней мере, шесть раз в течение каждого цикла выращивания.Были убраны только два внутренних ряда, а урожайность зерна измерялась в тоннах с гектара при влажности зерна 12,5%.
Дистанционное зондирование и обработка данных
Полеты выполнялись с использованием платформы RS eBee на базе БПЛА (SenseFly Ltd., Cheseaux-Lausanne, Швейцария) весом <2 кг, включая камеру и аккумулятор. Номинальная дальность радиосвязи платформы составляла 3 км при максимальном времени полета ~ 30–35 мин, крейсерской скорости 11–25 м / с и сопротивлении ветру до 12 м / с.БПЛА был оснащен мультиспектральной камерой MultiSpec 4C, которая обеспечивала спектральные изображения при 550 (полная ширина 40 нм на полувысоте, FWHM), 660 (40 нм FWHM), 735 (10 нм FWHM) и 790 нм (40 нм FWHM). ) и тепловизионной инфракрасной камеры ThermoMAP (7,5–13,5 мкм) (Айринов, Париж, Франция). Две камеры были установлены отдельно, и последовательные полеты проводились разными камерами. БПЛА пролетел 55 м над землей в полдень в солнечных условиях, охватив площадь 1,7 га (т. Е. Площадь на 0,2 га больше экспериментальной площади, чтобы получить точные ортофотопланы).Изображения были получены с 80% боковым и 90% продольным перекрытием, при полетах на север / юг и восток / запад. Это привело к разрешению на местности 6 и 12 см для мультиспектральной и тепловизионной камер соответственно. Для мультиспектральной камеры радиометрические калибровки и корректировки выполнялись перед каждым полетом с использованием стандартной панели камеры, предоставленной производителем. Кроме того, во время каждого полета яркость солнечного света измерялась датчиком падающего света, встроенным в мультиспектральную камеру, что позволяло проводить радиометрическую корректировку изображений, снятых в различных условиях освещения.Полеты и оценка визуальных заболеваний проводились в одни и те же дни, начиная с цветения. Четыре и три полета были выполнены в 2016 и 2017 годах соответственно.
Изображения были помечены геотегами для обработки ортофотоплана с использованием программного обеспечения Pix4D Mapper ® (v3.3.24; Pix4D, Лозанна, Швейцария). Изображения были преобразованы в коэффициент отражения и температуру поверхности для мультиспектральных и радиометрических данных теплового инфракрасного излучения соответственно. Всего для каждого ортофотоплана было рассчитано восемь различных вегетационных индексов (ВИ).Структурные VI представляли собой: нормализованный разностный вегетационный индекс (NDVI), перенормированный DVI (RDVI), оптимизированный вегетационный индекс с поправкой на почву (OSAVI), модифицированное простое соотношение (MSR) и модифицированное поглощение хлорофилла в индексах отражения (MCARI1 и MCARI2), в то время как пигмент конкретное простое соотношение для хлорофилла A (PSSRa) использовалось в качестве индекса, связанного с хлорофиллом, а зеленый цвет (G) использовался как индекс соотношения красный-зеленый-синий (RGB) (Таблица 2). Для каждой длины волны, необходимой для расчета VI, учитывалась ближайшая длина волны многоспектрального сигнала, принимая во внимание FWHM каждого канала.Температура купола оценивалась по тепловому инфракрасному сигналу.
Таблица 2 . Взаимосвязь между урожайностью зерна гибридов кукурузы и площадью под кривой прогрессирования заболевания (AUDPC) комплекса смоляных пятен с площадями под разными длинами волн, вегетативными индексами и тепловизионными изображениями при фунгицидных и нефунгицидных обработках.
Данные изображения были извлечены только из двух центральных рядов (из четырех рядов) каждого участка, так как только они были собраны для оценки урожайности зерна.Были очерчены два многоугольника (рис. 2В), по одному для каждого центрального ряда. Площадь 0,2 × 0,5 м, окружающая два центральных ряда, считалась буферной зоной. Пиксели были выбраны и усреднены изнутри полигонов с использованием программного обеспечения ArcGIS ® (v10.1; ESRI, Redlands, CA, USA).
Анализ данных
Дисперсионный анализ (ANOVA) был проведен, чтобы определить, одинаково ли работали генотипы кукурузы между экспериментальными годами с точки зрения снижения урожайности зерна, вызванного TSC.Фенотипические данные анализировали с помощью стандартной линейной смешанной модели, в которой год, репликация и графики в пределах репликации рассматривались как случайные эффекты. Обработки (фунгицидные и нефунгицидные) и генотипы считались фиксированными эффектами. Данные о заболевании, полученные с помощью визуальной оценки в двух циклах, были обобщены и проанализированы с использованием площади под кривой прогрессирования заболевания (AUDPC) трапециевидным методом, то есть интегралами Римана (Vanderplank, 1963). Площадь под кривой (AUC) для отдельных длин волн, вегетативных индексов (VI) и тепловых изображений также была получена с использованием интегралов Римана.Это позволяет интегрировать временную информацию из данных изображений и измерений заболевания в отдельные переменные (AUDPC и AUC). Различия в показателях индивидуального генотипа кукурузы оценивали по потерям урожая зерна (т / га) для каждой посадки при двух обработках. Данные AUDPC и AUC были использованы для индивидуальной ассоциации посредством регрессионного анализа с урожайностью зерна и друг с другом. Анализы проводились с использованием статистического программного обеспечения R, версия 3.3.3 и соответствующих библиотек для смешанных моделей и процедур множественного сравнения (Bates et al., 2015; Lenth, 2016; R Core Team, 2017).
Результаты
Влияние комплекса смоляных пятен на урожайность зерна
Результаты ANOVA показали, что влияние взаимодействия экспериментальных лет с генотипами кукурузы было незначительным ( p > 0,05, данные не показаны). Таким образом, эффект TSC был проанализирован в течение обоих экспериментальных лет. Развитие заболевания было оптимальным в течение обоих циклов, при этом чувствительные проверки достигли наивысших значений AUDPC среди всех генотипов за 2 года (Таблица 1).Борьба с болезнями при обработке фунгицидами также была оптимальной, и только следы болезни наблюдались на нижней части листвы, что приводило к низким показателям AUDPC. Это объяснило сильную взаимосвязь между урожайностью и AUDPC (Таблица 2) при нефунгицидной обработке ( R 2 = 0,84). Как и ожидалось, связь между AUDPC и урожаем зерна при обработке фунгицидами была слабой ( R 2 = 0,14).
Сравнение эффективности отдельных генотипов при двух обработках (фунгицид vs.нефунгициды) показали, что урожайность большинства генотипов была затронута (снижена) болезнью при обработке нефунгицидами (Таблица 1). Единственными исключениями были CLTHW13001 и Resistant Check 1, выходы которых существенно не различались ( p > 0,05) между двумя обработками. Наибольшая потеря урожая — 58% — наблюдалась на ПС-464.
Дистанционное зондирование
Анализ взаимосвязи урожайности зерна с AUC для отдельных длин волн, VI, температуры растительного покрова и AUDPC за 2 года показал, что большинство переменных RS сильно коррелировали с урожайностью при обработке нефунгицидами ( р ≤ 0.01, таблица 2). При нефунгицидной обработке коэффициент детерминации ( R 2 ) взаимодействий между урожаем зерна и отдельными длинами волн (550, 660, 735 и 790 нм) составлял 0,40–0,79. R 2 отношения структурных VI (NDVI, RDVI, OSAVI, MSR, MCARI1 и MCARI2) составляло 0,55–0,81. Кроме того, R 2 взаимосвязи индекса хлорофилла PSSRa и отношения RGB G с урожаем зерна было равно 0.79 и 0,76 соответственно. Эти данные означают, что генотипы с более высокими значениями AUC для этих переменных, за исключением W660, также имели более высокие урожаи в условиях болезни. Напротив, при нефунгицидной обработке значения температуры растительного покрова R 2 и AUDPC составляли 0,81 и 0,84 соответственно. Это указывало на то, что генотипы с более низкими значениями AUC температуры растительного покрова и более низкими показателями AUDPC имели более высокие урожаи в условиях болезни. В то время как отношения для различных длин волн, ИВ, температуры растительного покрова и AUDPC с урожайностью зерна были заметно выше при обработке нефунгицидами, эти отношения были предсказуемо слабее при обработке фунгицидами.Это объясняется отсутствием болезни, то есть фотосинтез и, следовательно, урожай зерна у здоровых растений не пострадали. При обработке фунгицидом связь W550, W660, W735, MSR и G с урожайностью была незначительной ( p > 0,05), однако W790 нм, другие показатели и температура растительного покрова имели значительную ( p ≤ 0,01) связь с доходностью. Среди всех переменных RS MCARI2 и температура растительного покрова имели самую сильную связь с урожайностью зерна ( R 2 = 0.81 для обоих) под давлением болезни (Таблица 2). Кроме того, MCARI1, MCARI2 и RDVI показали самые сильные связи со значениями AUDPC ( R 2 = 0,93 для каждого индекса).
Обсуждение
Наши результаты показывают, что потенциальные потери урожая от TSC у гибридов кукурузы могут достигать 58% у чувствительных генотипов в условиях сильного воздействия болезни. Это число значительно выше, чем 46%, о которых сообщает Bajet et al. (1994). Хотя влияние болезни на инбредные линии в нашем исследовании не исследовалось, потери урожая зерна также, вероятно, будут значительными.Сильная взаимосвязь между AUDPC и урожаем зерна ( R 2 = 0,84) указывает на общее влияние TSC на производство кукурузы после серьезной эпидемии в восприимчивой зародышевой плазме кукурузы. Тяжесть заражения TSC на станции Агуа Фриа в течение двух лет, в течение которых проводился эксперимент, не была особенно сильной, поэтому наши результаты представляют собой консервативную оценку потери урожая. Более серьезные природные эпидемии, которые случаются время от времени, могут привести к еще большим потерям урожая зерна.
Начало болезни в зависимости от стадии роста кукурузы (например, симптомы болезни, появляющиеся до или после цветения) может иметь большое влияние на потери урожая, хотя эта гипотеза требует дальнейшего тестирования. Еще одним фактором, влияющим на потери урожая, может быть роль M. maydis в эпидемиологии TSC. Hock et al. (1989) предположили, что M. maydis может продуцировать токсины, вызывающие ускоренное старение на листве кукурузы. В отдельных исследованиях (Loladze et al., неопубликованные данные), выявили наличие таких фитотоксинов в нескольких изолятах M. maydis . Различия в продукции фитотоксина между изолятами, возможно, предполагают существование разных рас в популяциях M. maydis . В зависимости от свойств продуцирования фитотоксинов у рас, присутствующих в конкретном месте и году, серьезность заболевания и степень повреждения, вызванного TSC, могут значительно варьироваться, что объясняет ежегодные колебания потерь урожая кукурузы.В настоящее время проводится исследование для проверки этой теории.
Дополнительным косвенным фактором, влияющим на потери урожая, вызванные TSC, могут быть гнили, которые часто обнаруживаются у растений, ослабленных TSC (Loladze et al., Неопубликованные данные). Хотя гнили не вызываются непосредственно теми же патогенами, которые вызывают TSC, они могут играть значительную роль в общей динамике потерь урожая. Взаимодействие между TSC и гнилью ушей, следовательно, также требует отдельного исследования.
Применение гиперспектральных сигналов для фенотипирования устойчивости к болезням ряда сельскохозяйственных культур широко обсуждалось Shakoor et al.(2017) и Simko et al. (2017). Примеры таких болезней включают полосатую ржавчину и фузариоз пшеницы (Bauriegel et al., 2011a; Devadas et al., 2015), апельсиновую ржавчину сахарного тростника (Apan et al., 2004), Venturia inaequalis на яблонях (Delalieux et al. ., 2007) и красное пятно на листьях миндаля (López-López et al., 2016). Во всех вышеупомянутых исследованиях сообщается о высокой точности гиперспектральных сигналов при применении к фенотипированию устойчивости к болезням.
Результаты текущего исследования, однако, демонстрируют возможность использования мультиспектральных изображений для оценки болезней кукурузы, метода, который значительно дешевле, чем использование гиперспектральных камер.Несколько предыдущих исследований показали, что NDVI, структурный индекс, рассчитываемый на основе длин волн красного и ближнего инфракрасного диапазона, был от умеренной до высокоточной при различении различных уровней серьезности болезней пшеницы и насекомых-вредителей. К ним относятся листовая ржавчина и полосовая ржавчина пшеницы, а также солнечный вредитель (Franke, Menz, 2007; Genc et al., 2008; Pretorius et al., 2017). Однако в нашем исследовании NDVI не был самым точным показателем с точки зрения корреляции с потерями урожая зерна кукурузы, вызванными TSC ( R 2 = 0.76).
Наше исследование показало, что количество ИВ, рассчитанных на основе многоспектрального сигнала и тепловых данных, сильно коррелировало с серьезностью заболевания и урожайностью зерна при нефунгицидных обработках. Самая сильная связь с урожайностью наблюдалась для MCARI2 VI и температуры растительного покрова ( R 2 = 0,81 для каждого, таблица 2). Это, однако, было слабее, чем связь между AUDPC и урожаем зерна ( R 2 = 0,84). Кроме того, сильная связь между MCARI2 и AUDPC ( R 2 = 0.93) предполагает, что этот индекс потенциально может служить вспомогательным инструментом для крупномасштабных исследований болезней, особенно на платформах высокопроизводительного фенотипирования. Хотя взаимосвязь между урожайностью зерна и AUDPC все еще была немного сильнее при обработке нефунгицидами ( R 2 = 0,84), как температуру растительного покрова, так и MCARI2 можно потенциально использовать для оценки устойчивости к болезням и, возможно, прогнозирования потерь урожая, вызванных TSC. в кукурузе. Следовательно, взаимосвязь между AUDPC и урожаем зерна все еще была немного выше, чем между AUC и урожаем зерна.Тем не менее, RS все еще имеет потенциальное применение для фенотипирования заболеваний на крупномасштабных платформах фенотипирования с высокой пропускной способностью. В таких случаях визуальная оценка отнимает слишком много времени.
VI MCARI2 — это улучшенная версия MCARI, которая была модифицирована с целью снижения шумового воздействия почвы в пределах сигнала отражения, сохраняя при этом чувствительность к индексу площади листьев кроны (LAI) и устойчивость к изменчивости содержания хлорофилла (Haboudane et al., 2004 г.).Ранее сообщалось, что область спектра, связанная со структурами навеса и листьев (красный край), была достаточно чувствительной, чтобы дифференцировать различные уровни тяжести заболевания (Garcia-Ruiz et al., 2013; Franceschini et al., 2019). Аналогичные результаты были получены в нашем исследовании, где красный край и NIR (735 и 790 нм) показали сильную взаимосвязь с урожайностью и AUDPC при нефунгицидных обработках (Таблица 2). Однако эти длины волн все еще имели несколько меньшую взаимосвязь с выходом и AUDPC по сравнению с MCARI2.
Растительность имеет два пика поглощения света в синей и красной областях спектра из-за содержания хлорофилла (C a + b ), высокой отражательной способности в зеленом цвете, в то время как ее биомасса и структура растительного покрова связаны с отражательной способностью в красном и ближнем ИК-спектрах. регионах (Richardson et al., 2002). ВП используются для объединения мультиспектральных наблюдений в единые метрики, которые сводят к минимуму влияние внешних факторов на спектральные данные и определяют конкретные характеристики полога (Baret and Guyot, 1991).Эти факты и возможное снижение LAI, вызванное TSC-индуцированным снижением фотосинтетической активности, привели нас к мысли, что MCARI2 также может использоваться в качестве потенциальной альтернативы для оценки потерь урожая, вызванных заболеванием.
В некоторых случаях развитие болезней растений может быть связано с изменениями температуры листвы больных растений, которые можно измерить с помощью инфракрасной термографии (Simko et al., 2017). Некоторые болезни растений могут вызывать закрытие устьиц, что снижает охлаждение за счет испарения и повышает температуру растительного покрова (Chaerle et al., 2001; Кальдерон и др., 2015). В текущем исследовании, хотя наблюдались заметные различия в температуре растительного покрова между устойчивыми и восприимчивыми генотипами, не было обнаружено прогрессирующей кривой температуры растительного покрова, параллельной кривой развития болезни (данные не показаны). Тем не менее, когда данные о температуре растительного покрова использовались для расчета AUC, такая информация временных рядов показала сильную взаимосвязь с данными урожайности при нефунгицидной обработке ( R 2 = 0.81). На отсутствие кривой роста температуры растительного покрова могли повлиять полевые условия (например, температура окружающей среды, тепловое излучение или интенсивность солнечного света) во время полетов и / или стадия развития растения и его взаимодействие с патогеном ( Chaerle et al., 1999, 2004; Lindenthal et al., 2005; Simko et al., 2017). В то же время следует отметить, что наши экспериментальные участки не подвергались водному стрессу или дефициту питательных веществ — факторам, которые могли повлиять на эвапотранспирацию и, следовательно, на температуру растительного покрова.
При условии, что абиотические стрессы, такие как засуха и нехватка питательных веществ в почве, сведены к минимуму во время испытаний болезней, RS может играть важную роль в высокопроизводительном фенотипировании устойчивости к заболеваниям листьев кукурузы. Это потенциально может снизить вероятность человеческой ошибки и снизить нагрузку на визуальную оценку в больших масштабах (Mahlein, 2016). С постоянным совершенствованием технологии RS и методов и процедур анализа данных изображений взаимосвязь между характеристиками растений и данными изображений может быть еще больше улучшена.Это может привести к более широкому и более распространенному применению технологии RS в селекции кукурузы на крупномасштабных платформах фенотипирования в разных местах.
В то время как технология RS имеет потенциал для значительных инноваций в высокопроизводительном фенотипировании устойчивости к болезням, барьеры, ограничивающие внедрение технологии в селекционные программы, остаются. Крупные начальные вложения в покупку системы, высококвалифицированные специалисты по сбору и обработке изображений, а также возможные задержки в обработке данных — вот некоторые из проблем (Khaled et al., 2017). Хотя технологии RS, которые включают камеры, платформы и программное обеспечение для обработки данных, становятся все более доступными, системы и протоколы необходимо адаптировать к требованиям фенотипирования конкретных, экономически важных болезней кукурузы.
Использование RS для ранней оценки воздействия болезни и выявления взаимодействий биотических и абиотических стрессов требует дальнейшего исследования, как это также было предложено предыдущими авторами (Calderón et al., 2013; Mahlein, 2016; Khaled et al., 2017; Симко и др., 2017). В связи с постоянным развитием технологии RS, возможности предсимптоматического раннего выявления болезней растений, как это было недавно сделано для оливковых культур, еще предстоит изучить на кукурузе (Zarco-Tejada et al., 2018). Это потенциально может помочь бороться с болезнями кукурузы с помощью соответствующих мероприятий по борьбе с болезнями до развития серьезных эпидемий.
Авторские взносы
AL разработал и провел полевое исследование и отвечал за компонент патологии растений.FR разработала и провела компонент исследования «Дистанционное зондирование». AL и FR внесли равный вклад в проект. FT провела статистический анализ. ФС предоставила зародышевую плазму и внесла свой вклад в рукопись. BG и MB предоставили критическую информацию о написании рукописи.
Финансирование
Эта работа была поддержана Исследовательской программой КГМСХИ по кукурузе (MAIZE). Содержание и мнения, выраженные здесь, принадлежат авторам и не обязательно отражают точку зрения авторского учреждения и не должны использоваться в целях рекламы или одобрения продукта.
Заявление о конфликте интересов
Авторы заявляют, что исследование проводилось при отсутствии каких-либо коммерческих или финансовых отношений, которые могут быть истолкованы как потенциальный конфликт интересов.
Благодарности
Авторы благодарят г-на Карлоса Муньоса и г-жу Лорену Гонсалес за их технический вклад в исследование. Мы благодарим г-жу Дженнифер Джонсон за рецензирование рукописи и Editage (www.editage.com) за редактирование на английском языке.
Список литературы
Альбетис, Дж., Жакен, А., Гулар, М., Пуилве, Х., Руссо, Дж., Клене, Х. и др. (2019). О возможностях мультиспектральных изображений БПЛА для обнаружения Flavescence dorée и болезней ствола виноградной лозы. Дистанционный датчик . 11:23. DOI: 10.3390 / RS11010023
CrossRef Полный текст | Google Scholar
Апан А., Хелд А., Финн С. и Маркли Дж. (2004). Обнаружение болезни сахарного тростника «оранжевая ржавчина» с помощью гиперспектральных изображений EO-1 Hyperion. Внутр. Дж. Дистанционный датчик . 25, 489–498. DOI: 10.1080 / 01431160310001618031
CrossRef Полный текст | Google Scholar
Бажет, Н. Б., Ренфро, Б. Л., и Карраско, Дж. М. В. (1994). Контроль дегтеобразующего пятна кукурузы и его влияние на урожайность. Внутр. Дж. Пешт Манаг . 40, 121–125. DOI: 10.1080 / 09670879409371868
CrossRef Полный текст | Google Scholar
Барет Ф. и Гайо Г. (1991). Возможности и пределы вегетационных индексов для оценки LAI и APAR. Среда удаленного сенсора . 35, 161–173. DOI: 10.1016 / 0034-4257 (91) -U
CrossRef Полный текст | Google Scholar
Бейтс Д., Мехлер М., Болкер Б. М. и Уокер С. (2015). Подгонка линейных моделей со смешанными эффектами с использованием lme4. J. Stat. Программное обеспечение . 67, 1–48. DOI: 10.18637 / jss.v067.i01
CrossRef Полный текст | Google Scholar
Бауригель Э., Гибель А. и Гейер М. (2011a). Раннее обнаружение инфекции фузариозом у пшеницы с помощью гиперспектральной визуализации. Comput. Электрон . Agric. 75, 304–312. DOI: 10.1016 / j.compag.2010.12.006
CrossRef Полный текст | Google Scholar
Bauriegel, E., Giebel, A., and Herppich, W. B. (2011b). Гиперспектральная визуализация и флюоресценция хлорофилла для анализа воздействия fusarium culmorum на фотосинтетическую целостность инфицированных колосьев пшеницы. Датчики 11, 3765–3779. DOI: 10.3390 / s110403765
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Bergsträsser, S., Fanourakis, D., Schmittgen, S., Cendrero-Mateo, M., Jansen, M., Scharr, H., et al. (2015). HyperART: неинвазивная количественная оценка характеристик листьев с использованием гиперспектральной визуализации поглощения-отражения-пропускания. Заводские методы 11, 1. doi: 10.1186 / s13007-015-0043-0
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Биссоннетт, С. (2015). Оповещение о болезни кукурузы: в 3 округах Северного Иллинойса выявлено новое грибковое заболевание листьев «дегтярное пятно» Phyllachora Maydist.Бык. Вредитель Манаг. Crop Dev. Инф. Иллинойс. Деп. Crop Sci. Унив . Доступно в Интернете по адресу: http://bulletin.ipm.illinois.edu/?p=3423 (по состоянию на 16 ноября 2017 г.).
Блэкберн Г. А. (1998). Спектральные индексы для оценки концентраций фотосинтетических пигментов: тест с использованием стареющих листьев деревьев. Внутр. Дж. Дистанционный датчик . 19, 657–675. DOI: 10.1080 / 014311698215919
CrossRef Полный текст | Google Scholar
Бок К. Х., Паркер П. Э., Кук А. З. и Готвальд Т.Р. (2008). Визуальная оценка и использование анализа изображений для оценки различных симптомов язвы цитрусовых на листьях грейпфрута. Завод Dis . 92, 530–541. DOI: 10.1094 / PDIS-92-4-0530
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Бок К. Х., Пул Г. Х., Паркер П. Э. и Готвальд Т. Р. (2010). Тяжесть болезней растений оценивается визуально, с помощью цифровой фотографии и анализа изображений, а также с помощью гиперспектральной визуализации. CRC. Крит. Rev. Plant Sci .29, 59–107. DOI: 10.1080 / 07352681003617285
CrossRef Полный текст | Google Scholar
Кальдерон, Р., Навас-Кортес, Дж. А., Лусена, К., и Зарко-Техада, П. Дж. (2013). Гиперспектральные и тепловые изображения с высоким разрешением для раннего обнаружения вертициллезного увядания оливок с использованием флуоресценции, температуры и узкополосных спектральных показателей. Remote Sens. Environ. 139, 231–245. DOI: 10.1016 / j.rse.2013.07.031
CrossRef Полный текст | Google Scholar
Кальдерон, Р., Навас-Кортес, Дж. А., и Зарко-Техада, П. Дж. (2015). Раннее обнаружение и количественная оценка вертициллезного увядания у оливок с использованием гиперспектральных и тепловых изображений на больших площадях. Remote Sens. 7, 5584–5610. DOI: 10.3390 / RS70505584
CrossRef Полный текст | Google Scholar
Себальос, Х., Дойч, Дж. А. (1992). Наследование устойчивости к комплексу смоляных пятен у кукурузы. Фитопатология 82, 505–512. DOI: 10.1094 / Phyto-82-505
CrossRef Полный текст | Google Scholar
Chaerle, L., Де Бовер, Ф., Ван Монтегю, М., и Ван дер Стретен, Д. (2001). Термографическая визуализация гибели клеток табака и арабидопсиса. Среда растительных клеток . 24, 15–25. DOI: 10.1046 / j.1365-3040.2001.00654.x
CrossRef Полный текст | Google Scholar
Chaerle, L., Hagenbeek, D., De Bruyne, E., Valck, R., and Van Der Straeten, D. (2004). Тепловая визуализация и визуализация флюоресценции хлорофилла позволяют выявить взаимодействия между растениями и патогенами на ранней стадии. Физиология растительных клеток .45, 887–896. DOI: 10.1093 / pcp / pch097
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Chaerle, L., Van Caeneghem, W., Messens, E., Lambers, H., Van Montagu, M., and Van Der Straeten, D. (1999). Пресимптоматическая визуализация взаимодействий растений с вирусами с помощью термографии. Нац. Биотехнология . 17, 813–816. DOI: 10.1038 / 11765
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Чен, Дж. М. (1996). Оценка вегетационных индексов и модифицированный простой коэффициент для бореальных применений. Банка. J. Remote Sens. 22, 229–242. DOI: 10.1080 / 07038992.1996.10855178
CrossRef Полный текст | Google Scholar
Delalieux, S., van Aardt, J., Keulemans, W., Schrevens, E., and Coppin, P. (2007). Обнаружение биотического стресса (Venturia inaequalis) у яблонь с использованием гиперспектральных данных: непараметрические статистические подходы и физиологические последствия. евро. J. Agron. 27, 130–143. DOI: 10.1016 / j.eja.2007.02.005
CrossRef Полный текст | Google Scholar
Девадас, Р., Лэмб Д. В., Бэкхаус Д. и Симпфендорфер С. (2015). Последовательное применение гиперспектральных индексов для выявления заражения полосатой ржавчиной и дефицита азота у пшеницы. Precis. Agric. 16, 477–491. DOI: 10.1007 / s11119-015-9390-0
CrossRef Полный текст | Google Scholar
Диттрих, У., Хок, Дж., Кранц, Дж., И Ренфро, Б. Л. (1991). Прорастание аскоспор Phyllachora maydis и конидий Monographella maydis. Криптогам. Бот. 2, 214–218.
Google Scholar
Franceschini, M.H.D, Bartholomeus, H., van Apeldoorn, D.F., Suomalainen, J., and Kooistra, L. (2019). Возможность получения оптических изображений с беспилотных летательных аппаратов для раннего выявления и оценки степени тяжести фитофтороза картофеля. Дистанционный датчик 11: 224. DOI: 10.3390 / RS11030224
CrossRef Полный текст | Google Scholar
Franke, J., and Menz, G. (2007). Выявление разновременных болезней пшеницы с помощью многоспектрального дистанционного зондирования. Precis. Agric. 8, 161–172. DOI: 10.1007 / s11119-007-9036-y
CrossRef Полный текст | Google Scholar
Гарсия-Руис, Ф., Шанкаран, С., Майя, Дж. М., Ли, В. С., Расмуссен, Дж., И Эхсани, Р. (2013). Сравнение двух платформ для получения изображений с воздуха для идентификации цитрусовых деревьев, зараженных Хуанлунбин. Comput. Электрон. Agric. 91, 106–115. DOI: 10.1016 / j.compag.2012.12.002
CrossRef Полный текст | Google Scholar
Genc, H., Genc, L., Турхан, Х., Смит, С., и Нэйшн, Дж. (2008). Индексы вегетации как индикаторы повреждения солнечными вредителями (Hemiptera: Scutelleridae) полевой пшеницы. Afr. J. Biotechnol. 7, 173–180. Доступно в Интернете по адресу: https://academicjournals.org/journal/AJB/article-abstract/9E68C996947
Google Scholar
Хабудан, Д., Миллер, Дж. Р., Патти, Э., Зарко-Техада, П. Дж., И Страчан, И. Б. (2004). Индексы гиперспектральной растительности и новые алгоритмы прогнозирования зеленого LAI растительного покрова: моделирование и проверка в контексте точного земледелия. Remote Sens. Environ. 90, 337–352. DOI: 10.1016 / j.rse.2003.12.013
CrossRef Полный текст | Google Scholar
Хансен, Дж., Дэвисон, Д., Джонс, Д., и Сан, X. (2016). Phyllachora Maydis, Пятно кукурузной смолы. Предупреждение о вредителях, Bur. Энтомол. Нематол. Завод Патол . Доступно в Интернете по адресу: http://www.freshfromflorida.com/content/download/69885/1624208/Pest_Alert_Phyllachora_maydis,_Corn_Tar_Spot.pdf (по состоянию на 15 ноября 2017 г.).
Хок, Дж. (1988). Таргетинг на пятнистую смолу 1987 Годовой отчет: Международный центр улучшения кукурузы и пшеницы (Мехико: CIMMYT).
Google Scholar
Хок, Дж., Диттрих, У., Ренфро, Б. Л., и Кранц, Дж. (1992). Последовательное развитие патогенов в комплексе болезни кукурузы. Mycopathologia 117, 157–161. DOI: 10.1007 / BF00442777
CrossRef Полный текст | Google Scholar
Хок, Дж., Кранц, Дж. И Ренфо, Б. Л. (1995). Исследования по эпидемиологии комплекса болезни дегтярных пятен кукурузы в Мексике. Завод Патол . 44, 490–502. DOI: 10.1111 / j.1365-3059.1995.tb01671.x
CrossRef Полный текст | Google Scholar
Хок, Дж., Кранц, Дж., И Ренфро, Б. (1989). El Complejo ‘mancha de asfalto’ de maíz: Su distribucción geográfica, Requisitos ambientales e importancia económica en México. Rev. Mex. Фитопатол. 7, 129–135.
Google Scholar
Халед А. Ю., Абд Азиз С., Беджо С. К., Нави Н. М., Семан И. А. и Онвуде Д. И. (2017). Раннее обнаружение заболеваний растительной ткани с помощью спектроскопии — области применения и ограничения. заявл. Spectrosc. Ред. . 53, 36–64. DOI: 10.1080 / 05704928.2017.1352510
CrossRef Полный текст | Google Scholar
Kuska, M., Wahabzada, M., Leucker, M., Dehne, H.-W., Kersting, K., Oerke, E.-C., et al. (2015). Гиперспектральное фенотипирование в микроскопическом масштабе: к автоматизированной характеристике взаимодействий между растениями и патогенами. Заводские методы 11:28. DOI: 10.1186 / s13007-015-0073-7
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Линденталь, М., Steiner, U., Dehne, H., and Oerke, E.-C. (2005). Влияние развития ложной мучнистой росы на транспирацию листьев огурца, визуализированное с помощью цифровой инфракрасной термографии. Фитопатология. 95, 233–240. DOI: 10.1094 / PHYTO-95-0233
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Лопес-Лопес, М., Кальдерон, Р., Гонсалес-Дуго, В., Зарко-Техада, Дж. П., и Феререс, Э. (2016). Раннее обнаружение и количественная оценка пятен на красных листах миндаля с использованием гиперспектральных и тепловых изображений с высоким разрешением. Дистанционный датчик . 8: 276. DOI: 10.3390 / RS8040276
CrossRef Полный текст | Google Scholar
Mahlein, A.-K. (2016). Обнаружение болезней растений с помощью датчиков изображения — параллели и особые требования для точного земледелия и фенотипирования растений. Завод Dis . 100, 241–251. DOI: 10.1094 / PDIS-03-15-0340-FE
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Mahlein, A.-K., Kuska, M. T., Thomas, S., Bohnenkamp, D., Alisaac, E., Behmann, J., и другие. (2017). Обнаружение болезней растений с помощью гиперспектральных изображений: от лаборатории до поля. Adv. Anim. Biosci. 8, 238–243. DOI: 10.1017 / S2040470017001248
CrossRef Полный текст | Google Scholar
Mahlein, A.-K., Steiner, U., Hillnhütter, C., Dehne, H.-W., and Oerke, E.-C. (2012). Гиперспектральная визуализация для мелкомасштабного анализа симптомов, вызванных различными заболеваниями сахарной свеклы. Методы растений 8: 3. DOI: 10.1186 / 1746-4811-8-3
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Мотталеб, К.А., Лоладзе А., Сондер К., Круземан Г. и Сан-Висенте Ф. (2018). Угрозы комплексного заболевания кукурузы, вызванного смоляными пятнами, в Соединенных Штатах Америки и его глобальные последствия. Mitig. Адаптировать. Strateg. Glob. Чанг . 24, 281–300. DOI: 10.1007 / s11027-018-9812-1
CrossRef Полный текст | Google Scholar
Ньютон, А.С., и Хакетт, К.А. (1994). Субъективные компоненты оценки милдью ярового ячменя. евро. J. Plant Pathol. 100, 395–412. DOI: 10.1007 / BF01874807
CrossRef Полный текст | Google Scholar
Наттер, Ф.У. младший (1993). Оценка точности, повторяемости внутри эксперта и надежности систем оценки заболеваний. Фитопатология 83, 806–812.
Google Scholar
Паркер С. Р., Шоу М. В. и Ройл Д. Дж. (1995). Достоверность визуальных оценок тяжести заболевания на листьях злаков. Завод Патол . 44, 856–864. DOI: 10.1111 / j.1365-3059.1995.tb02745.x
CrossRef Полный текст | Google Scholar
Преториус, З.А., Lan, C.X., Prins, R., Knight, V., McLaren, N.W., Singh, R.P. и др. (2017). Применение дистанционного зондирования для определения локусов устойчивости взрослых растений к полосатой ржавчине в двух картированных популяциях мягкой пшеницы. Precis. Сельское хозяйство . 18, 411–428. DOI: 10.1007 / s11119-016-9461-x
CrossRef Полный текст | Google Scholar
R Основная команда (2017). R: язык и среда для статистических вычислений . Вена: Фонд R для статистических вычислений.
PubMed Аннотация
Ричардсон, А.Д., Дуйган, С. П., и Берлин, Г. П. (2002). Оценка неинвазивных методов оценки содержания хлорофилла в листьях. Н. Фитол . 153, 185–194. DOI: 10.1046 / j.0028-646X.2001.00289.x
CrossRef Полный текст | Google Scholar
Рондо, Г., Стивен, М., и Барет, Ф. (1996). Оптимизация вегетационных индексов с поправкой на почву. Remote Sens. Environ. 55, 95–107. DOI: 10.1016 / 0034-4257 (95) 00186-7
CrossRef Полный текст | Google Scholar
Ружан, Дж.Л. и Бреон Ф. М. (1995). Оценка PAR, поглощенного растительностью, на основе двунаправленных измерений коэффициента отражения. Remote Sens. Environ. 51, 375–384. DOI: 10.1016 / 0034-4257 (94) 00114-3
CrossRef Полный текст | Google Scholar
Роуз, Дж. У., Хаас, Р. Х., Шелл, Дж. А., и Диринг, Д. У. (1973). Мониторинг весеннего наступления и ретроградации (эффект зеленой волны) естественной растительности . Прог. Rep. RSC 1978-1.
Руль, Г., Ромберг, М.К., Биссоннетт, С., Плева, Д., Кресвелл, Т., и Уайз, К.А. (2016). Первое сообщение о пятнах смолы на кукурузе, вызванных Phyllachora maydis в США. Завод Дис. 100: 1496. DOI: 10.1094 / PDIS-12-15-1506-PDN
CrossRef Полный текст | Google Scholar
Шакур, Н., Ли, С., и Моклер, Т. К. (2017). Высокопроизводительное фенотипирование для ускорения селекции сельскохозяйственных культур и мониторинга болезней в поле. Curr. Opin. Завод Биол . 38, 184–192. DOI: 10.1016 / j.pbi.2017.05.006
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Стеддом К., Бредехофт М. В., Хан М. и Раш К. М. (2005). Сравнение визуальной и мультиспектральной радиометрической оценки болезни пятнистости листьев Cercospora сахарной свеклы. Завод Дис. 89, 153–158. DOI: 10.1094 / PD-89-0153
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Вандерпланк, Дж. Э. (1963). Болезни растений: эпидемии и борьба .Нью-Йорк, Нью-Йорк: Academic Press, 349.
Янг К., Эверитт Дж. Х. и Фернандес К. Дж. (2010). Сравнение мультиспектральных и гиперспектральных изображений, полученных с воздуха, для картирования корневой гнили хлопчатника. Биосист. Eng . 107, 131–139. DOI: 10.1016 / j.biosystemseng.2010.07.011
CrossRef Полный текст | Google Scholar
Зарко-Техада, П. Дж., Берхон, А., Лопес-Лозано, Р., Миллер, Дж. Р., Мартин, П., Качорро, В. и др. (2005). Оценка состояния виноградника с помощью гиперспектральных индексов: моделирование отражательной способности листьев и кроны в прерывистом кроне со структурой рядов. Среда удаленного сенсора . 99, 271–287. DOI: 10.1016 / j.rse.2005.09.002
CrossRef Полный текст | Google Scholar
Зарко-Техада, П. Дж., Камино, К., Бек, П. С. А., Кальдерон, Р., Хорнеро, А., Эрнандес-Клементе, Р. и др. (2018). Превизуальные симптомы инфекции Xylella fastidiosa проявляются в спектральных изменениях признаков растений. Нац. Растения 4, 432–439. DOI: 10.1038 / s41477-018-0189-7
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Упражнения с отягощениями инициируют механическую мишень транслокации рапамицина (mTOR) и локализацию белкового комплекса в скелетных мышцах человека
Бурд, Н. А., Танг, Дж. Э., Мур, Д. Р. и Филлипс, С. М. Тренировка с упражнениями и метаболизм белка: влияние сокращения, потребления белка и различий по полу. Журнал прикладной физиологии 106 , 1692–1701, DOI: 10.1152 / japplphysiol.
.2008 (2009).
CAS Статья PubMed Google Scholar
Laplante, M. & Sabatini, D. Передача сигналов M. mTOR в контроле роста и болезни. Ячейка 149 , 274–293, DOI: 10.1016 / j.cell.2012.03.017 (2012).
CAS Статья PubMed PubMed Central Google Scholar
Betz, C. & Hall, M. N. Где находится mTOR и что он там делает? Журнал клеточной биологии 203 , 563–574, DOI: 10.1083 / jcb.201306041 (2013).
CAS Статья PubMed PubMed Central Google Scholar
Дикинсон, Дж. М. и др. . Активация комплекса 1 рапамицина у млекопитающих необходима для стимуляции синтеза белка скелетных мышц человека незаменимыми аминокислотами. Журнал питания 141 , 856–862, DOI: 10.3945 / jn.111.139485 (2011).
CAS Статья PubMed PubMed Central Google Scholar
Драммонд, М. Дж. и др. .Введение рапамицина людям блокирует вызванное сокращением увеличение синтеза белка скелетных мышц. Журнал физиологии 587 , 1535–1546, DOI: 10.1113 / jphysiol.2008.163816 (2009).
CAS Статья PubMed PubMed Central Google Scholar
МакГлори, С. и Филлипс, С. М. Оценка регуляции пластичности скелетных мышц в ответ на прием белка и упражнения с отягощениями: последние разработки. Текущее мнение в области клинического питания и метаболической помощи 17 , 412–417, DOI: 10.1097 / MCO.0000000000000083 (2014).
Артикул PubMed Google Scholar
Биоло, Г., Магги, С. П., Уильямс, Б. Д., Типтон, К. Д. и Вулф, Р. Р. Увеличение скорости обмена мышечного белка и транспорта аминокислот после упражнений с отягощениями у людей. Американский физиологический журнал 268 , E514–520 (1995).
CAS PubMed Google Scholar
Филлипс, С. М., Типтон, К. Д., Арсланд, А., Вольф, С. Э. и Вулф, Р. Р. Синтез и распад смешанного мышечного белка после упражнений с отягощениями у людей. Американский физиологический журнал 273 , E99–107 (1997).
CAS PubMed Google Scholar
Элиассон, Дж. и др. . Максимальные удлиняющие сокращения увеличивают фосфорилирование киназы p70 S6 в скелетных мышцах человека при отсутствии питания. Американский физиологический журнал. Эндокринология и метаболизм 291 , E1197–1205, DOI: 10.1152 / ajpendo.00141.2006 (2006).
CAS Статья PubMed Google Scholar
Burd, N.A. et al. . Повышенная аминокислотная чувствительность при синтезе миофибриллярного белка сохраняется до 24 часов после упражнений с отягощениями у молодых мужчин. Журнал питания 141 , 568–573, DOI: 10.3945 / jn.110.135038 (2011).
CAS Статья PubMed Google Scholar
Биоло, Г., Типтон, К. Д., Кляйн, С. и Вулф, Р. Р. Обильный запас аминокислот усиливает метаболический эффект упражнений на мышечный белок. Американский физиологический журнал 273 , E122–129 (1997).
CAS PubMed Google Scholar
Борсхайм, Э., Типтон, К. Д., Вольф, С. Э. и Вулф, Р. Р. Восстановление незаменимых аминокислот и мышечного белка после упражнений с отягощениями. Американский физиологический журнал. Эндокринология и метаболизм 283 , E648–657, DOI: 10.1152 / ajpendo.00466.2001 (2002).
CAS Статья PubMed Google Scholar
Мур, Д. Р., Атертон, П. Дж., Ренни, М. Дж., Тарнопольский, М. А. и Филлипс, С. М. Упражнения с отягощениями усиливают передачу сигналов mTOR и MAPK в мышцах человека по сравнению с тем, что наблюдается в состоянии покоя после приема болюсного белка. Acta Physiol (Oxf) 201 , 365–372, DOI: 10.1111 / j.1748-1716.2010.02187.x (2011).
CAS Статья Google Scholar
Дрейер, Х. К. и др. .Прием обогащенных лейцином незаменимых аминокислот и углеводов после упражнений с отягощениями усиливает передачу сигналов mTOR и синтез белка в мышцах человека. Американский физиологический журнал. Эндокринология и метаболизм 294 , E392–400, DOI: 10.1152 / ajpendo.00582.2007 (2008).
CAS Статья PubMed Google Scholar
Sancak, Y. et al. . Комплекс Ragulator-Rag направляет mTORC1 на поверхность лизосом и необходим для его активации аминокислотами. Ячейка 141 , 290–303, DOI: 10.1016 / j.cell.2010.02.024 (2010).
CAS Статья PubMed PubMed Central Google Scholar
Inoki, K., Li, Y., Xu, T. и Guan, K. L. Rheb GTPase является прямой мишенью для активности TSC2 GAP и регулирует передачу сигналов mTOR. Гены и развитие 17 , 1829–1834, DOI: 10.1101 / gad.1110003 (2003).
CAS Статья Google Scholar
Manning, BD, Tee, AR, Logsdon, MN, Blenis, J. & Cantley, LC Идентификация продукта гена супрессора опухоли комплекса-2 туберозного склероза туберин в качестве мишени фосфоинозитид-3-киназы / akt путь. Молекулярная ячейка 10 , 151–162 (2002).
CAS Статья PubMed Google Scholar
Tee, A.R., Manning, B.D., Roux, P.P., Cantley, L.C., Blenis, J. Продукты гена комплекса туберозного склероза, Tuberin и Hamartin, контролируют передачу сигналов mTOR, действуя как комплекс белка, активирующего GTPase, по отношению к Rheb. Текущая биология: CB 13 , 1259–1268 (2003).
CAS Статья PubMed Google Scholar
Jacobs, B. L. et al. . Эксцентрические сокращения увеличивают фосфорилирование комплекса туберозного склероза-2 (TSC2) и изменяют нацеливание TSC2 и механистическую мишень рапамицина на лизосомы. Журнал физиологии 591 , 4611–4620, DOI: 10.1113 / jphysiol.2013.256339 (2013).
CAS Статья PubMed PubMed Central Google Scholar
Якобс, Б. Л., Гудман, К. А. и Хорнбергер, Т. А. Механическая активация передачи сигналов mTOR: новая роль для нацеливания на поздние эндосомы / лизосомы. Журнал исследований мышц и подвижности клеток 35 , 11–21, DOI: 10.1007 / s10974-013-9367-4 (2014).
CAS Статья PubMed Google Scholar
Risson, V. et al. . Мышечная инактивация mTOR вызывает метаболические и дистрофиновые дефекты, приводящие к тяжелой миопатии. Журнал клеточной биологии 187 , 859–874, DOI: 10.1083 / jcb.2001 (2009).
CAS Статья PubMed PubMed Central Google Scholar
Джейкобс, Б. Л. и др. . Идентификация механически регулируемых сайтов фосфорилирования на туберине (TSC2), которые контролируют механистическую мишень передачи сигналов рапамицина (mTOR). Журнал биологической химии . DOI: 10.1074 / jbc.M117.777805 (2017).
Google Scholar
Bodine, S. C. et al. . Путь Akt / mTOR является важным регулятором гипертрофии скелетных мышц и может предотвратить атрофию мышц in vivo . Природа клеточной биологии 3 , 1014–1019, DOI: 10.1038 / ncb1101-1014 (2001).
CAS Статья PubMed Google Scholar
Филп А., Гамильтон Д. Л. и Баар К. Сигналы, опосредующие ремоделирование скелетных мышц с помощью упражнений с отягощениями: независимая от PI3-киназы активация mTORC1. J Appl Physiol 110 , 561–568, DOI: 10.1152 / japplphysiol.00941.2010 (2011).
CAS Статья PubMed Google Scholar
Мур, Д. Р. и др. . Дифференциальная стимуляция синтеза миофибриллярных и саркоплазматических белков при приеме белков в покое и после упражнений с отягощениями. Журнал физиологии 587 , 897–904, DOI: 10.1113 / jphysiol.2008.164087 (2009).
CAS Статья PubMed PubMed Central Google Scholar
Staples, A. W. et al. . Углеводы не увеличивают индуцированное физическими упражнениями накопление белка по сравнению с одним белком. Медицина и наука в спорте и физических упражнениях 43 , 1154–1161, DOI: 10.1249 / MSS.0b013e31820751cb (2011).
CAS Статья PubMed Google Scholar
Корольчук В.И. и др. . Позиционирование лизосом координирует реакцию клеток на питательные вещества. Природа клеточной биологии 13 , 453–460, DOI: 10,1038 / ncb2204 (2011).
CAS Статья PubMed PubMed Central Google Scholar
Fujita, S., Rasmussen, B. B., Bell, J. A., Cadenas, J. G. & Volpi, E. Кинетика внутриклеточных аминокислот базальных мышц у женщин и мужчин. Американский физиологический журнал. Эндокринология и метаболизм 292 , E77–83, DOI: 10.1152 / ajpendo.00173.2006 (2007).
CAS Статья PubMed Google Scholar
Habets, P.E. et al. . Содержание РНК различается в медленных и быстрых мышечных волокнах: значение для интерпретации изменений экспрессии мышечных генов. Журнал гистохимии и цитохимии: официальный журнал Общества гистохимиков 47 , 995–1004 (1999).
CAS Статья Google Scholar
Хорн, З. и Хескет, Дж. Иммунологическая локализация рибосом в поперечно-полосатых мышцах крысы. Доказательства миофибриллярной ассоциации и онтологических изменений в субарколемме: миофибриллярное распределение. Биохимический журнал 268 , 231–236 (1990).
CAS Статья PubMed PubMed Central Google Scholar
Horne, Z. & Hesketh, J. Повышенная ассоциация рибосом с миофибриллами во время гипертрофии скелетных мышц, вызванной либо кленбутеролом, агонистом бета-адренорецепторов, либо тенотомией. Биохимический журнал 272 , 831–833 (1990).
CAS Статья PubMed PubMed Central Google Scholar
Гудман, К. А. и др. . Новое понимание регуляции синтеза белка в скелетных мышцах, выявленное с помощью нового нерадиоактивного метода in vivo . Журнал FASEB: официальное издание Федерации американских обществ экспериментальной биологии 25 , 1028–1039, DOI: 10.1096 / fj.10-168799 (2011).
CAS Статья Google Scholar
Гудман, К. А., Пьер, П. и Хорнбергер, Т. А. Визуализация синтеза белка с пуромицином. Труды Национальной академии наук Соединенных Штатов Америки 109 , E989; ответ автора E990, DOI: 10.1073 / pnas.1202000109 (2012).
Aspuria, P. J. & Tamanoi, F.Семейство Rheb GTP-связывающих белков. Сотовая сигнализация 16 , 1105–1112, DOI: 10.1016 / j.cellsig.2004.03.019 (2004).
CAS Статья PubMed Google Scholar
Ти, А. Р. и др. . Продукты гена комплекса-1 и -2 туберозного склероза функционируют вместе, подавляя у млекопитающих-мишень опосредованной рапамицином (mTOR) нижестоящей передачи сигналов. Труды Национальной академии наук Соединенных Штатов Америки 99 , 13571–13576, DOI: 10.1073 / pnas.202476899 (2002).
ADS CAS Статья PubMed PubMed Central Google Scholar
Long, X., Lin, Y., Ortiz-Vega, S., Yonezawa, K. & Avruch, J. Rheb связывает и регулирует киназу mTOR. Текущая биология: CB 15 , 702–713, DOI: 10.1016 / j.cub.2005.02.053 (2005).
CAS Статья PubMed Google Scholar
Long, X., Ortiz-Vega, S., Lin, Y. & Avruch, J. Связывание Rheb с мишенью рапамицина (mTOR) млекопитающих регулируется достаточностью аминокислот. Журнал биологической химии 280 , 23433–23436, DOI: 10.1074 / jbc.C500169200 (2005).
CAS Статья PubMed Google Scholar
Avruch, J. et al. . Активация mTORC1 в два этапа: активация каталитической функции Rheb-GTP и усиление связывания субстратов с хищником. Сделки Биохимического общества 37 , 223–226, DOI: 10.1042 / BST0370223 (2009).
CAS Статья PubMed Google Scholar
Inoki, K., Li, Y., Zhu, T., Wu, J. & Guan, K. L. TSC2 фосфорилируется и ингибируется Akt и подавляет передачу сигналов mTOR. Природа клеточной биологии 4 , 648–657, DOI: 10.1038 / ncb839 (2002).
CAS Статья PubMed Google Scholar
Greenhaff, P. L. et al. . Диссоциация между эффектами аминокислот и инсулина на передачу сигналов, убиквитинлигазы и обмен белка в мышцах человека. Американский физиологический журнал. Эндокринология и метаболизм 295 , E595–604, DOI: 10.1152 / ajpendo.
.2008 (2008).CAS Статья PubMed PubMed Central Google Scholar
You, J. S. et al. . Роль диацилглицеринкиназы дзета и фосфатидной кислоты в механической активации мишени рапамицина (mTOR) млекопитающих и гипертрофии скелетных мышц. Журнал биологической химии 289 , 1551–1563, DOI: 10.1074 / jbc.M113.531392 (2014).
CAS Статья PubMed Google Scholar
Harris, T. E. et al. .mTOR-зависимая стимуляция ассоциации eIF4G и eIF3 инсулином. Журнал EMBO 25 , 1659–1668, DOI: 10.1038 / sj.emboj.7601047 (2006).
CAS Статья PubMed PubMed Central Google Scholar
Holz, M. K., Ballif, B. A., Gygi, S. P. & Blenis, J. mTOR и S6K1 опосредуют сборку комплекса преинициации трансляции посредством динамического обмена белками и событий упорядоченного фосфорилирования. Ячейка 123 , 569–580, DOI: 10.1016 / j.cell.2005.10.024 (2005).
CAS Статья PubMed Google Scholar
Дрейер, Х. К. и др. . Упражнения с отягощениями увеличивают активность AMPK и снижают фосфорилирование 4E-BP1 и синтез белка в скелетных мышцах человека. Журнал физиологии 576 , 613–624, DOI: 10.1113 / jphysiol.2006.113175 (2006).
CAS Статья PubMed PubMed Central Google Scholar
McGlory, C. et al. . Применение анализа [гамма-32P] АТФ-киназы для изучения анаболической передачи сигналов в скелетных мышцах человека. Журнал прикладной физиологии 116 , 504–513, DOI: 10.1152 / japplphysiol.01072.2013 (2014).
CAS Статья PubMed PubMed Central Google Scholar
Бергстром, Дж. Чрескожная пункционная биопсия скелетных мышц в физиологических и клинических исследованиях. Скандинавский журнал клинических и лабораторных исследований 35 , 609–616 (1975).
CAS Статья PubMed Google Scholar
Burd, N.A. et al. . Упражнения с отягощениями с малой нагрузкой и большим объемом стимулируют синтез мышечного белка больше, чем упражнения с отягощениями с высокой нагрузкой и низким объемом, у молодых мужчин. PloS one 5 , e12033, DOI: 10.1371 / journal.pone.0012033 (2010).
ADS Статья PubMed PubMed Central Google Scholar
Мур, Д. Р. и др. . Дозовая реакция потребляемого белка в мышцах и синтез белка альбумина после упражнений с отягощениями у молодых мужчин. Американский журнал клинического питания 89 , 161–168, DOI: 10.3945 / ajcn.2008.26401 (2009).
CAS Статья PubMed Google Scholar
Комплексные импедансы, краткое введение
Комплексные импедансы, краткое введениеКомплексные импедансы
Вернуться к оглавлению.
С помощью следующего калькулятора несколько свойств рассчитываются для ряда
схема строится с резистором и катушкой или конденсатором.
Введите значение частоты, резистора и катушки / конденсатора желтым цветом.
поля и щелкните
«вычислить».
Ниже объясняется, что вычисляет этот калькулятор.
Что такое импеданс
Импеданс — это значение сопротивления, которое кажется электрическому компоненту.
иметь с определенной частотой.
Импеданс обозначается буквой X, единица измерения
Ом (Ом).
Для резистора это полное сопротивление: X = R
Для катушки это полное сопротивление: X = 2.pi.f.L
Для конденсатора это полное сопротивление: X = 1 / (2.pi.f.C)
X = полное сопротивление (Ом)
pi = 3.1415
f = частота (Герц)
L = индукция катушки (Генри)
C = емкость конденсатора (Фарад)
Однако этот способ описания импеданса не является полным, поскольку
фаза между напряжением и током не показана.
По значению X мы не можем понять, резистор ли это, катушка или конденсатор.
Комплексный импеданс
Комплексный импеданс создается действительной частью (R = резистор) последовательно с
мнимая часть (+ JX = катушка или -JX = конденсатор).
Комплексный импеданс обозначается буквой Z, а единица измерения
Ω.
Обозначение комплексного импеданса может быть Z = R + JX.
В этом случае резистор и катушка соединены последовательно.
Импеданс катушки: X = 2.pi.f.L
В катушке (переменное) напряжение всегда опережает ток на 90,
это обозначено + J.
Обозначение комплексного импеданса также может быть: Z = R-JX.
В этом случае резистор и конденсатор соединены последовательно.
Импеданс конденсатора X = 1 / (2.pi.f.C)
С конденсатором (переменное) напряжение всегда будет на 90 ниже
текущий, это обозначается -J.
Пример 1: Z1 = 220 + J300 Ом.
В этом примере резистор 220 Ом
и катушка с сопротивлением 300 Ом
соединены последовательно.
Эти два последовательно соединенных компонента составляют один комплексный импеданс.
Пример 2: Z2 = 470-J80 Ом
В этом примере резистор 470 Ом
и конденсатор с сопротивлением 80 Ом
соединены последовательно.
Пример 3: Z3 = 100 + J0 Ом
Это чистый резистор 100 Ом (при этом
частота).
Поскольку мнимая часть равна нулю, мы также можем написать: Z3 = 100-J0
Ом
Пример 4: Z4 = 0 + J60
Ω
Это катушка с сопротивлением 60 Ом, у этой катушки нет последовательного соединения.
сопротивление.
Пример 5: Z5 = 0-J400 Ом
Это конденсатор с импедансом 400
Ω, этот конденсатор не имеет последовательного сопротивления.
Оператор J
Буква J в комплексных импедансах называется оператором J.
В резисторе напряжение на резисторе и ток через него равны
по фазе нет разности фаз.
Импеданс резистора называется реальным сопротивлением.
Импеданс катушки не действительный, а мнимый.
В катушке напряжение всегда опережает ток на 90, это отображается
+ J, за которым следует значение импеданса.
Конденсатор — это тоже мнимое сопротивление.
В конденсаторе напряжение всегда на 90 ниже силы тока, это отображается
с -J, за которым следует значение импеданса.
Расчет с помощью оператора J
Если мы проводим расчет с мнимым импедансом, применяются следующие правила:
Дж = √-1
Дж = -1
1 / Дж = -Дж
1 / -J =
Дж.Ja + Jb = J (a + b)
J-a = -Ja
Суммирование комплексных сопротивлений
Если два комплексных импеданса соединены последовательно, новый комплексный импеданс
сформирован.
При сложении двух комплексных импедансов мы можем сложить реальные части, а также добавить
мнимые части.
Пример: Z1 и Z2 соединены последовательно, сумма этих двух равна Z6.
Z1 = 220 + J300 Ом
Z2 = 470-J80 Ом +
Z6 = 690 + J220 Ом
Мнимые части складываются, но поскольку мнимая часть Z2 равна
отрицательный, он фактически вычитается из мнимой части Z1.
Другой пример: Z7 = Z1 + Z2 + Z3 + Z4 + Z5
Z1 = 220 + J300 Ом.
Z2 = 470-J80 Ом
Z3 = 100 + J0 Ом
Z4 = 0 + J60 Ом
Z5 = 0-J400 Ом +
Z7 = 790-J120 Ом
Сумма всех этих импедансов ведет себя на этой частоте так же, как
резистор 790 Ом последовательно с конденсатором с
Сопротивление 120 Ом.
Resonan ce
Если конденсатор и катушка соединены последовательно, а мнимые части равны,
они добавят к нулю Ω.
Тогда цепь находится в последовательном резонансе, и только сопротивление обоих
компонентов осталось.
При последовательном резонансе полное сопротивление контура LC достигает минимального значения.
стоимость.
При параллельных цепях LC полное сопротивление будет достичь максимального значения при резонансе.
Фактор Q
Мы можем вычислить добротность (Q) комплексного импеданса.
Отношение добротности между мнимой и действительной частями импеданса.
Q = X / R
Неважно, положительная или отрицательная мнимая часть в
при расчетах мы используем только число за J.
Q не имеет единицы измерения, и значение всегда положительное (или ноль, в случае чистого
резистор).
Пример: Z7 = 790-J120 Ом имеет добротность 0,1519
Абсолютное значение импеданса
Если мы подключим переменное напряжение к комплексному импедансу, ток будет
поток.
Чтобы вычислить значение тока, нам нужно знать абсолютное значение
сопротивление.
Абсолютное значение указано | Z | и единица
Ω.
Для комплексного импеданса Z = RJX абсолютное значение:
| Z | = √ (R + X)
Пример: Z7 = 790-J120 Ом
| Z7 | = √ (790 + 120) = 799 Ом.
Если этот импеданс подключен к переменному напряжению, ток будет течь с
значение:
I = U / | Z |
Пример: напряжение на Z7 составляет 10 В (среднеквадратичное значение).
| Z7 | = 799 Ом
I = 10/799 = 0,0125 Ампера (среднеквадратичное значение).
Фаза между напряжением и током
Фаза между напряжением на комплексном импедансе и током
через него можно рассчитать следующим образом:
Фаза = арктангенс (X / R).
Единица — градусы ()
Значение X может быть как положительным, так и отрицательным, в зависимости от знака перед
оператор J.
При положительном значении фазы напряжение опережает ток.
При отрицательном значении фазы напряжение отстает от тока.
Значение фазы может изменяться от +90 (катушка) через 0 (резистор) до -90 (конденсатор).
Пример:
С комплексным импедансом Z7 = 790-J120 Ом
фаза между напряжением и током:
Фаза = арктангенс (-120 / 790) = -8,6
Импедансы в виде векторов
Комплексные импедансы могут быть помещены в виде векторов на диаграмму.
Угол с горизонтальной осью указывает фазу между напряжением и тока, длина вектора соответствует значению импеданса.
| | |
| Комплексное сопротивление: Z8 = 4 + J3 Ом Длина вектора Z8 равна абсолютному значению | Z8 |. В данном случае: | Z8 | = √ (4 + 3) = 5 Ом | Два комплексных импеданса (Z8 и Z9) соединены последовательно. Z10 — это сумма Z8 и Z9, поэтому Z10 = Z8 + Z9 |
Преобразование последовательного импеданса в параллельный импеданс
Комплексный импеданс, состоящий из резистора, включенного последовательно с катушкой / конденсатором
может быть преобразован в параллельную цепь резистора и катушки / конденсатора.
Обе схемы будут вести себя одинаково на этой частоте, но это
Верно только для одной частоты, на которой мы рассчитываем схему.
| Последовательная цепь резистора и катушки преобразуется в параллельную цепь резистора и катушки. | |
| Последовательная цепь резистора и конденсатора преобразуется в параллельную цепь резистора и конденсатора. |
Преобразование работает следующим образом:
У нас есть сложная последовательная цепь Z = RS + JXS
RS и XS — это серия
составные части.
С помощью следующих формул мы можем найти значения для параллельных компонентов RP
ru XP.
Rp = (RS + XS) / RS
Xp = J (RS + XS) / XS
Когда комплексный импеданс является емкостным, поэтому Z = RS-JXS
тогда также значение XP будет отрицательным.
Пример 1:
Комплексное сопротивление Z = 20 + J15 Ом
Параллельные импедансы:
RP = (20 + 15) / 20 = 31,25 Ом
XP = J (20 + 15) / 15 =
+ J41,67 Ом
Последовательная цепь резистора и катушки преобразуется в параллельную цепь
резистор и катушка
Пример 2:
Те же значения компонентов, но теперь для емкостного импеданса
Комплексный импеданс Z = 20-Дж15 Ом
Параллельные импедансы:
RP = (20 + (- 15)) / 20 = 31,25
Ом
XP = Дж (20 + (- 15)) / — 15
= -J41.67 Ом
Последовательная цепь резистора и конденсатора преобразуется в параллельную.
резистора и конденсатора.
Вернуться к оглавлению.
Филогенетически информативные мутации в генах, влияющих на устойчивость к антибиотикам в комплексе Mycobacterium tuberculosis | Геномная медицина
Партнерство «Остановить туберкулез» | Лидеры G20 поднимают проблему туберкулеза до уровня глав государств [Интернет]. [цитируется 24 ноября 2017 г.]. Доступно по адресу: http://www.stoptb.org/news/stories/2017/ns17_044.жерех
Дхеда К., Гамбо Т., Маартенс Дж., Дули К.Э., Мюррей М., Фурин Дж. И др. Комиссия по респираторной медицине Lancet: новости 2019 г .: эпидемиология, патогенез, передача, диагностика и лечение неизлечимого туберкулеза с множественной лекарственной устойчивостью. Ланцет Респир Мед. 2019; 7: 820–6.
CAS PubMed PubMed Central Статья Google Scholar
Schön T, Miotto P, Köser CU, Viveiros M, Böttger E, Cambau E.Тестирование на лекарственную устойчивость Mycobacterium tuberculosis: проблемы, последние разработки и перспективы. Clin Microbiol Infect Off Publ Eur Soc Clin Microbiol Infect Dis. 2017; 23: 154–60.
Google Scholar
Heyckendorf J, Andres S, Köser CU, Olaru ID, Schön T, Sturegård E, et al. Что такое сопротивление? Влияние тестирования фенотипической и молекулярной лекарственной устойчивости на терапию туберкулеза с множественной и широкой лекарственной устойчивостью. Антимикробные агенты Chemother.2018; 62 (2).
Всемирная организация здравоохранения. (2018). Технический отчет о критических концентрациях для тестирования лекарственной чувствительности лекарственных средств, используемых для лечения лекарственно-устойчивого туберкулеза. Всемирная организация здоровья. https://apps.who.int/iris/handle/10665/260470. Лизенз: CC BY-NC-SA 3.0 IGO.
Miotto P, Tessema B, Tagliani E, Chindelevitch L, Starks AM, Emerson C, et al. Стандартизированный метод интерпретации связи между мутациями и фенотипической лекарственной устойчивостью Mycobacterium tuberculosis.Eur Respir J. 2017; 50 (6).
PubMed PubMed Central Статья CAS Google Scholar
Консорциум CRyPTIC и проект 100 000 геномов. Прогнозирование восприимчивости к противотуберкулезным препаратам первого ряда с помощью секвенирования ДНК. N Engl J Med. 2018; 379: 1403–15.
Артикул Google Scholar
Köser CU, Feuerriegel S, Summers DK, Archer JAC, Niemann S.Важность генетического разнообразия в составе комплекса Mycobacterium tuberculosis для разработки новых антибиотиков и диагностических тестов лекарственной устойчивости. Антимикробные агенты Chemother. 2012. 56 (12): 6080–7.
PubMed PubMed Central Статья CAS Google Scholar
Coll F, McNerney R, Guerra-Assunção JA, Glynn JR, Perdigão J, Viveiros M, et al. Надежный штрих-код SNP для типирования штаммов комплекса Mycobacterium tuberculosis.Nat Commun. 2014; 5: 4812.
CAS PubMed PubMed Central Статья Google Scholar
Feuerriegel S, Köser CU, Niemann S. Филогенетические полиморфизмы в генах устойчивости к антибиотикам комплекса Mycobacterium tuberculosis. J Antimicrob Chemother. 2014; 69: 1205–10.
CAS PubMed PubMed Central Статья Google Scholar
Эль Ачкар С., Деманче С., Осман М., Рафей Р., Исмаил М.Б., Яакуб Х. и др.Туберкулез с лекарственной устойчивостью, Ливан, 2016–2017 гг. Emerg Infect Dis 2019; 25: 564–568.
Ajileye A, Alvarez N, Merker M, Walker TM, Akter S, Brown K, et al. Некоторые синонимичные и несинонимичные мутации gyrA в Mycobacterium tuberculosis приводят к систематическим ложноположительным результатам анализа устойчивости к фторхинолонам с помощью тестов Hain GenoType MTBDRsl. Антимикробные агенты Chemother. 2017; 61 (4).
Глобальная лабораторная инициатива. Линейные зондовые тесты для выявления лекарственно-устойчивого туберкулеза.Руководство по интерпретации и отчетности для сотрудников лабораторий и врачей. http://www.stoptb.org/wg/gli/assets/documents/LPA_test_web_ready.pdf (дата обращения 2.11.2018).
Comas I, Coscolla M, Luo T., Borrell S, Holt KE, Kato-Maeda M, et al. Миграция за пределы Африки и неолитическое распространение Mycobacterium tuberculosis с современными людьми. Нат Жене. 2013; 45: 1176–82.
CAS PubMed PubMed Central Статья Google Scholar
Коль Т.А., Утпател С., Шлейзенер В., Де Филиппо М.Р., Бекерт П., Чирилло Д.М. и др. MTBseq: комплексный инструмент для анализа последовательности всего генома изолятов комплекса Mycobacterium tuberculosis. PeerJ. 2018; 6: e5895.
PubMed PubMed Central Статья CAS Google Scholar
Li H, Durbin R. Быстрое и точное согласование коротких считываний с помощью преобразования Барроуза-Уиллера. Bioinforma Oxf Engl. 2009; 25: 1754–60.
CAS Статья Google Scholar
Маккенна А., Ханна М., Бэнкс Е., Сиваченко А., Цибульскис К., Керницкий А. и др. Набор инструментов для анализа генома: платформа MapReduce для анализа данных секвенирования ДНК следующего поколения. Genome Res. 2010; 20: 1297–303.
CAS PubMed PubMed Central Статья Google Scholar
Li H, Handsaker B, Wysoker A, Fennell T, Ruan J, Homer N, et al. Формат выравнивания / карты последовательностей и SAMtools. Bioinforma Oxf Engl. 2009; 25: 2078–9.
Артикул CAS Google Scholar
Цена МН, Дехал П.С., Аркин А.П. FastTree 2 — деревья приблизительно максимального правдоподобия для больших трасс. PLoS One. 2010; 5: e9490.
PubMed PubMed Central Статья CAS Google Scholar
FigTree [Интернет]. [цитировано 23 октября 2013 г.]. Доступно по адресу: http://tree.bio.ed.ac.uk/software/figtree/.
Merker M, Blin C, Mona S, Duforet-Frebourg N, Lecher S, Willery E, et al. Эволюционная история и глобальное распространение линии Пекинской микобактерии туберкулеза. Нат Жене. 2015; 47: 242–9.
CAS PubMed PubMed Central Статья Google Scholar
Brosch R, Gordon SV, Marmiesse M, Brodin P, Buchrieser C, Eiglmeier K, et al. Новый сценарий эволюции комплекса Mycobacterium tuberculosis. Proc Natl Acad Sci U S A.2002; 99: 3684–9.
CAS PubMed PubMed Central Статья Google Scholar
Walker TM, Merker M, Knoblauch AM, Helbling P, Schoch OD, van der Werf MJ, et al. Кластер микобактерий туберкулеза с множественной лекарственной устойчивостью среди пациентов, прибывающих в Европу с Африканского Рога: молекулярно-эпидемиологическое исследование. Lancet Infect Dis. 2018.
Lee AS, Teo AS, Wong SY. Новые мутации в ndh в изолятах микобактерий туберкулеза, устойчивых к изониазиду.Антимикробные агенты Chemother. 2001; 45: 2157–9.
CAS PubMed PubMed Central Статья Google Scholar
Brossier F, Veziris N, Truffot-Pernot C, Jarlier V, Sougakoff W. Молекулярное исследование устойчивости к противотуберкулезному препарату этионамид в клинических изолятах Mycobacterium tuberculosis с множественной лекарственной устойчивостью. Антимикробные агенты Chemother. 2011; 55: 355–60.
CAS PubMed PubMed Central Статья Google Scholar
Machado D, Perdigão J, Ramos J, Couto I, Portugal I, Ritter C и др. Высокий уровень устойчивости к изониазиду и этионамиду у микобактерий Mycobacterium tuberculosis семейства Lisboa с множественной лекарственной устойчивостью связан с двойными мутациями inhA. J Antimicrob Chemother. 2013; 68: 1728–32.
CAS PubMed PubMed Central Статья Google Scholar
Морлок ГП, Метчок Б., Сайкс Д., Кроуфорд Дж. Т., Кукси Р. Локусы ethA, inhA и katG клинических изолятов микобактерий туберкулеза, устойчивых к этионамиду.Антимикробные агенты Chemother. 2003; 47: 3799–805.
CAS PubMed PubMed Central Статья Google Scholar
Xu J, Wang B, Hu M, Huo F, Guo S, Jing W и др. Первичная устойчивость к клофазимину и бедаквилину среди изолятов от больных туберкулезом с множественной лекарственной устойчивостью. Антимикробные агенты Chemother. 2017; 61 (6).
Desjardins CA, Cohen KA, Munsamy V, Abeel T., Maharaj K, Walker BJ, et al.Геномный и функциональный анализы штаммов Mycobacterium tuberculosis указывают на участие ald в устойчивости к D-циклосерину. Нат Жене. 2016; 48: 544–51.
CAS PubMed PubMed Central Статья Google Scholar
Чен Дж. М., Уплекар С., Гордон С. В., Коул СТ. Точечная мутация в cycA частично способствует признаку устойчивости к D-циклосерину вакцинных штаммов Mycobacterium bovis BCG. PLoS One. 2012; 7: e43467.
CAS PubMed PubMed Central Статья Google Scholar
Andries K, Villellas C, Coeck N, Thys K, Gevers T., Vranckx L, et al. Приобретенная устойчивость Mycobacterium tuberculosis к бедаквилину. PLoS One. 2014; 9: e102135.
PubMed PubMed Central Статья CAS Google Scholar
Ривз А.З., Кэмпбелл П.Дж., Султана Р., Малик С., Мюррей М., Пликайтис Б.Б. и др. Перекрестная резистентность к аминогликозидам у Mycobacterium tuberculosis из-за мутаций в 5′-нетранслируемой области whiB7.Антимикробные агенты Chemother. 2013; 57: 1857–65.
CAS PubMed PubMed Central Статья Google Scholar
Köser CU, Bryant JM, Parkhill J, Peacock SJ. Последствия мутации whiB7 (Rv3197A) в изолятах генотипа Beijing из комплекса Mycobacterium tuberculosis. Антимикробные агенты Chemother. 2013; 57: 3461.
PubMed PubMed Central Статья CAS Google Scholar
Накатани И., Опель-Ридинг Х.К., Меркер М., Мачадо Д., Андрес С., Кумар С.С. и др. Роль мутаций аланинрацемазы в устойчивости Mycobacterium tuberculosis к D-циклосерину. Антимикробные агенты Chemother. 2017; 61 (12).
Китинг Л.А., Уиллер П.Р., Мансур Х., Инвальд Дж. К., Дейл Дж., Хьюинсон Р. Г. и др. Потребность в пирувате некоторых членов комплекса Mycobacterium tuberculosis обусловлена неактивной пируваткиназой: последствия для роста in vivo. Mol Microbiol.2005; 56: 163–74.
CAS PubMed PubMed Central Статья Google Scholar
Риохас М.А., Макгоф К.Дж., Райдер-Риохас С.Дж., Растоги Н., Хазбон М.Х. Филогеномный анализ видов комплекса Mycobacterium tuberculosis показывает, что Mycobacterium africanum, Mycobacterium bovis, Mycobacterium caprae, Mycobacterium microti и Mycobacterium pinnipedii являются более поздними гетеротипическими синонимами Mycobacterium tuberculosis.Int J Syst Evol Microbiol. 2018; 68: 324–32.
CAS PubMed PubMed Central Статья Google Scholar
Ates LS, Dippenaar A, Sayes F, Pawlik A, Bouchier C, Ma L, et al. Неожиданное геномное и фенотипическое разнообразие Mycobacterium africanum линии 5 влияет на лекарственную устойчивость, секрецию белка и иммуногенность. Genome Biol Evol. 2018; 10: 1858–74.
CAS PubMed PubMed Central Статья Google Scholar
Marmiesse M, Brodin P, Buchrieser C, Gutierrez C, Simoes N, Vincent V и др. Макромассив и биоинформатический анализ выявляют микобактериальные «основные» гены, вариации в семействе генов ESAT-6 и новые филогенетические маркеры комплекса Mycobacterium tuberculosis. Microbiol Read Engl. 2004; 150: 483–96.
CAS Статья Google Scholar
Abdallah AM, Hill-Cawthorne GA, Otto TD, Coll F, Guerra-Assunção JA, Gao G, et al.Каталог геномной экспрессии глобальной коллекции вакцинных штаммов БЦЖ свидетельствует о сильно различающихся адаптациях метаболизма и клеточной стенки. Научный доклад 2015; 5: 15443.
CAS PubMed PubMed Central Статья Google Scholar
Yew WW, Liang D, Chan DP, Shi W, Zhang Y. Молекулярные механизмы устойчивости к клофазимину у Mycobacterium tuberculosis. J Antimicrob Chemother. 2017; 72: 2943–4.
CAS PubMed PubMed Central Статья Google Scholar
Zhang S, Chen J, Cui P, Shi W., Zhang W., Zhang Y. Идентификация новых мутаций, связанных с устойчивостью к клофазимину у Mycobacterium tuberculosis. J Antimicrob Chemother. 2015; 70: 2507–10.
CAS PubMed PubMed Central Статья Google Scholar
Ахтман М. Эволюция, популяционная структура и филогеография генетически мономорфных бактериальных патогенов. Annu Rev Microbiol. 2008; 62: 53–70.
CAS PubMed PubMed Central Статья Google Scholar
McDERMOTT W, Tompsett R. Активация пиразинамида и никотинамида в кислой среде in vitro. Am Rev Tuberc. 1954; 70: 748–54.
CAS PubMed PubMed Central Google Scholar
Луазо С., Менардо Ф., Асеффа А., Хайлу Е., Гуми Б., Амени Дж. И др. Африканское происхождение Mycobacterium bovis. Evol Med Public Health. [цитировано 19 февраля 2020 г.]; Доступно по ссылке: https://academic.oup.com/emph/advance-article/doi/10.1093/emph/eoaa005/5719036.
Луазо С., Бритес Д., Мозер И., Колл Ф., Пурсель С., Робб-Остерман С. и др. Пересмотренная интерпретация Hain Lifescience GenoType MTBC для дифференциации Mycobacterium canettii и членов комплекса Mycobacterium tuberculosis. Антимикробные агенты Chemother. 2019; 63 (6).
Müller B, Dürr S, Alonso S, Hattendorf J, Laisse CJM, Parsons SDC, et al. Зоонозный туберкулез человека, вызванный Mycobacterium bovis. Emerg Infect Dis. 2013; 19: 899–908.
PubMed PubMed Central Статья Google Scholar
Feuerriegel S, Köser CU, Baù D, Rüsch-Gerdes S, Summers DK, Archer JAC, et al. Влияние разнообразия Fgd1 и ddn в комплексе Mycobacterium tuberculosis на чувствительность к PA-824 in vitro. Антимикробные агенты Chemother. 2011; 55: 5718–22.
CAS PubMed PubMed Central Статья Google Scholar
Feuerriegel S, Köser CU, Richter E, Niemann S. Mycobacterium canettii по своей природе устойчива как к пиразинамиду, так и к пиразиноевой кислоте. J Antimicrob Chemother. 2013; 68: 1439–40.
CAS PubMed PubMed Central Статья Google Scholar
Deshpande D, Alffenaar J-WC, Köser CU, Dheda K, Chapagain ML, Simbar N, et al. Фармакокинетика / фармакодинамика, восприимчивость и дозировка D-циклосерина при туберкулезе с множественной лекарственной устойчивостью: дело Фауста.Clin Infect Dis Off Publ Infect Dis Soc Am. 2018; 67: S308–16.
CAS Статья Google Scholar
Кезер К.Ю., Маурер Ф.П., Кранцер К. «Те, кто не может вспомнить прошлое, обречены его повторить»: тесты на лекарственную чувствительность к бедаквилину и деламаниду. Int J Infect Dis IJID Off Publ Int Soc Infect Dis. 2019; 80С: S32–5.
Google Scholar
Шен Т., Матушек Э., Мохамед С., Утукури М., Хейселл С., Алффенаар Дж. В. и др.Стандарты тестирования MIC, которые применяются к большинству бактериальных патогенов, также должны быть внедрены в отношении Mycobacterium tuberculosis complex. Clin Microbiol Infect Off Publ Eur Soc Clin Microbiol Infect Dis. 2019; 25: 403–5.
Google Scholar
EUCAST. СОП для калибровки методов суррогатной МИК для M. tuberculosis по сравнению с эталонным методом МИК EUCAST. 2019.
Google Scholar
Köser CU, Javid B, Liddell K, Ellington MJ, Feuerriegel S, Niemann S, et al. Механизмы лекарственной устойчивости и противотуберкулезные препараты. Lancet Lond Engl. 2015; 385: 305–7.
Артикул Google Scholar
Кальметер Г. Лекция Гаррода в 2014 году: EUCAST — движемся ли мы к международному соглашению? J Antimicrob Chemother. 2015; 70: 2427–39.
PubMed PubMed Central Статья CAS Google Scholar
Kahlmeter G, Giske CG, Kirn TJ, Sharp SE. Точка-контрапункт: различия между Европейским комитетом по тестированию чувствительности к противомикробным препаратам и рекомендациями Института клинических лабораторных стандартов по сообщению результатов определения чувствительности к противомикробным препаратам. J Clin Microbiol. 2019; 57 (9).
Gumbo T, Lenaerts AJ, Hanna D, Romero K, Nuermberger E. Доклинические модели разработки противотуберкулезных препаратов: анализ ландшафта. J Infect Dis. 2015; 211 (Приложение 3): S83–95.
PubMed PubMed Central Статья Google Scholar
Произошла ошибка при настройке пользовательского файла cookie
Этот сайт использует файлы cookie для повышения производительности. Если ваш браузер не принимает файлы cookie, вы не можете просматривать этот сайт.
Настройка вашего браузера для приема файлов cookie
Существует множество причин, по которым cookie не может быть установлен правильно. Ниже приведены наиболее частые причины:
- В вашем браузере отключены файлы cookie.Вам необходимо сбросить настройки своего браузера, чтобы он принимал файлы cookie, или чтобы спросить вас, хотите ли вы принимать файлы cookie.
- Ваш браузер спрашивает вас, хотите ли вы принимать файлы cookie, и вы отказались. Чтобы принять файлы cookie с этого сайта, нажмите кнопку «Назад» и примите файлы cookie.
- Ваш браузер не поддерживает файлы cookie. Если вы подозреваете это, попробуйте другой браузер.
- Дата на вашем компьютере в прошлом. Если часы вашего компьютера показывают дату до 1 января 1970 г., браузер автоматически забудет файл cookie.Чтобы исправить это, установите правильное время и дату на своем компьютере.
- Вы установили приложение, которое отслеживает или блокирует установку файлов cookie. Вы должны отключить приложение при входе в систему или проконсультироваться с системным администратором.
Почему этому сайту требуются файлы cookie?
Этот сайт использует файлы cookie для повышения производительности, запоминая, что вы вошли в систему, когда переходите со страницы на страницу. Чтобы предоставить доступ без файлов cookie потребует, чтобы сайт создавал новый сеанс для каждой посещаемой страницы, что замедляет работу системы до неприемлемого уровня.
Что сохраняется в файле cookie?
Этот сайт не хранит ничего, кроме автоматически сгенерированного идентификатора сеанса в cookie; никакая другая информация не фиксируется.
Как правило, в файле cookie может храниться только информация, которую вы предоставляете, или выбор, который вы делаете при посещении веб-сайта. Например, сайт не может определить ваше имя электронной почты, пока вы не введете его. Разрешение веб-сайту создавать файлы cookie не дает этому или любому другому сайту доступа к остальной части вашего компьютера, и только сайт, который создал файл cookie, может его прочитать.
Индуцибельная и приобретенная устойчивость к кларитромицину в комплексе Mycobacterium abscessus
Аннотация
Назначение
Кларитромицин считался краеугольным камнем в лечении комплексных инфекций Mycobacterium abscessus . Были описаны механизмы генетической устойчивости, и многие эксперты предлагают амикацин в качестве альтернативы. Тем не менее кларитромицин имеет несколько преимуществ; следовательно, необходимо идентифицировать нефункциональный аллель erm (41) , чтобы определить наиболее подходящее лечение.Цели этого исследования состояли в том, чтобы охарактеризовать молекулярные механизмы устойчивости к кларитромицину в коллекции из комплексных изолятов Mycobacterium abscessus и проверить взаимосвязь между этими механизмами и антибиотикограммой.
Материалы и методы
Клинические изоляты М . abscessus комплекс (n = 22) у 16 пациентов был идентифицирован с использованием четырех генов домашнего хозяйства ( rpoB , secA1 , sodA и hsp65 ), и их генетическая устойчивость была охарактеризована путем изучения erm (41) и rrl генов.Девять штаммов были выделены из клинических изолятов и подвергнуты Е-тесту и тестам на чувствительность к кларитромицину с микроразведением с показаниями через 3, 7 и 14 дней.
Результаты
Мы классифицировали 11/16 (68,8%) M . абсцесс subsp. абсцесс , 4/16 (25,0%) М . абсцесс subsp. боллетий и 1/16 (6,3%) M . абсцесс subsp. massiliense . Аллель T28 erm (41) наблюдался у 8 Mycobacterium abscessus subps. abscessus и 3 Mycobacterium abscessus subsp. боллетий . Один штамм М . абсцесс subsp. bolletii имел усеченный ген erm (41) и был чувствителен к кларитромицину. В первых изолятах гена rrl мутаций не наблюдалось. У трех пациентов наблюдение за исходными штаммами rrl дикого типа показало приобретенную устойчивость.
Выводы
Большинство клинических изолятов M . abscessus комплекс обладал индуцируемой резистентностью к кларитромицину и полным отсутствием конститутивной резистентности. Наши результаты показали, что приобретение мутаций устойчивости в гене rrl было связано с функциональным и нефункциональным геном erm (41) . Необходимо соблюдать осторожность при использовании только секвенирования erm (41) для идентификации M . абсцесс подвид. В этом исследовании сообщается о приобретенной мутации в положении 2057 гена rrl , придающей конститутивную резистентность к кларитромицину от средней до низкой.
Образец цитирования: Rubio M, March F, Garrigó M, Moreno C, Español M, Coll P (2015) Индуцируемая и приобретенная устойчивость к кларитромицину в комплексе Mycobacterium abscessus . PLoS ONE 10 (10): e0140166. https://doi.org/10.1371/journal.pone.0140166
Редактор: Андрес Р. Флото, Кембриджский университет, СОЕДИНЕННОЕ КОРОЛЕВСТВО
Поступила: 11 февраля 2015 г .; Принята к печати: 17 августа 2015 г .; Опубликован: 8 октября 2015 г.
Авторские права: © 2015 Rubio et al.Это статья в открытом доступе, распространяемая в соответствии с условиями лицензии Creative Commons Attribution License, которая разрешает неограниченное использование, распространение и воспроизведение на любом носителе при условии указания автора и источника.
Доступность данных: Все соответствующие данные находятся в пределах бумага.
Финансирование: Эта работа была поддержана Министерством экономики и конкуренции, Instituto de Salud Carlos III и софинансирована Европейским региональным фондом развития, ЕФРР «Путь к достижению Европы» и Испанской сетью исследований. по инфекционным болезням (REIPI) RD12 / 0015/0017.Финансирующие организации не играли никакой роли в дизайне исследования, сборе и анализе данных, принятии решения о публикации или подготовке рукописи.
Конкурирующие интересы: Авторы заявили об отсутствии конкурирующих интересов.
Введение
Комплексные подвиды Mycobacterium abscessus представляют собой быстрорастущие микобактерии (RGM), вызывающие хронические легочные инфекции, кожные инфекции и, в некоторых случаях, бактериемию [1,2]. Эти подвиды считаются наиболее устойчивыми к лекарствам из всех RGM [3–5], что затрудняет лечение вызываемых ими инфекций.Макролиды, такие как кларитромицин и азитромицин, считались краеугольным камнем в лечении комплексных инфекций Mycobacterium abscessus [5]. Были описаны механизмы молекулярной устойчивости, и многие эксперты предлагают амикацин в качестве альтернативы [6]. Тем не менее кларитромицин имеет несколько преимуществ; следовательно, необходимо идентифицировать нефункциональный аллель erm (41) , чтобы определить наиболее подходящее лечение.
Устойчивость к кларитромицину в M . abscessus комплекс может быть конститутивным или индуцибельным, в зависимости от двух механизмов. Конститутивная резистентность включает спонтанные точечные мутации, выбранные во время химиотерапии на основе макролидов, в положениях 2058 и 2059 гена rrl , который кодирует 23S рРНК [7]. Второй механизм связан с индуцибельной устойчивостью [8], включающей присутствие функционального гена erm (41) и последующее метилирование рибосомы. Конститутивная резистентность характеризуется in vitro высокими значениями устойчивости к кларитромицину через 3 дня культивирования [4,7], в то время как индуцибельная резистентность, связанная с геном erm (41) , характеризуется только значениями устойчивости к МИК in vitro. в 7 и даже 14 дней [9].
Таксономия M . абсцесс Группа сложна и до сих пор не решена. В 2011 году М . боллетий и М . massiliense были отнесены к одному подвиду ( M . abscessus subsp. bolletii ) из-за их низкого генетического разнообразия [10]. В 2013 году анализ данных полногеномного секвенирования подтвердил дифференциацию M . abscessus сложен на 3 подвида: M . абсцесс subsp. абсцесс , М . абсцесс subsp. боллетий и М . абсцесс subsp. massiliense [11]. В настоящем исследовании мы используем эту номенклатуру для трех подвидов.
Широко признано, что для идентификации трех подвидов в комплексе необходимо несколько генов домашнего хозяйства [12,13]. Эти три подвида тесно связаны, рекомбинация и перенос генов описаны [11].Гены, наиболее часто используемые для классификации M . Подвиды abscessus представляют собой гены r poB , secA1 , sodA , recA и hsp65 [12,13]. Даже когда используются новые мощные методы, такие как матричная лазерная десорбция / ионизация по времени пролета (MALDI-TOF) [14], идентификация M . abscessus сложных изолятов на уровне подвидов не является 100% точным.
Ген erm (41) немного отличается между M . абсцесс сложный подвид. Полный ген erm (41) с 10 секвеварами описан для M . абсцесс subsp. abscessus [15]: секвевары с нуклеотидом Т28 связаны с индуцибельной устойчивостью к кларитромицину, а с нуклеотидом С28 связаны с чувствительностью к кларитромицину. В М . абсцесс subsp. bolletii , ген erm (41) аналогичен гену T28 M . абсцесс subsp. абсцесс [16]. В М . абсцесс subsp. massiliense , ген erm (41) , как известно, имеет 2 делеции, что делает его нефункциональным. Анализ последовательности гена erm (41) был использован для классификации M . abscessus сложный подвид [17].
Собрано М . abscessus комплекс клинических изолятов с 1995 по 2014 год в университетской больнице в Испании.Цели этого исследования состояли в том, чтобы охарактеризовать молекулярные механизмы устойчивости к кларитромицину в коллекции исходных изолятов комплекса Mycobacterium abscessus , идентифицированных на уровне подвидов, и проверить взаимосвязь между механизмами генетической устойчивости и результатами антибиотикограммы. Мы также изучили 10 изолятов при наблюдении за 4 пациентами с помощью молекулярного типирования и фенотипических и генотипических антибиотиков.
Материалы и методы
Заявление об этике
Это исследование было ретроспективным без взаимодействия с пациентами, и вся информация о пациентах была деидентифицирована перед анализом.Поэтому комитеты по этике Fundació de Gestió Sanitaria del Hospital de la Santa Creu i Sant Pau отказались от необходимости получать информированное согласие и одобрили это исследование (IISBP-CLA-2014-23).
Образцы
В период с 1995 по 2014 год было получено 22 клинических изолята от 16 пациентов: 3 — от кожных и 19 — от легочных инфекций. Для наблюдения использовали десять изолятов от 4 пациентов: 3 от пациента 1, 3 от пациента 2, 2 от пациента 3 и 2 от пациента 4. ДНК из всех изолятов была выделена для генотипической идентификации и анализа молекулярной устойчивости к кларитромицину.Девять первых выделенных изолятов от 9 отдельных пациентов были подвергнуты тестам на чувствительность к кларитромицину.
Классификация подвидов
Тотальную ДНК экстрагировали из клинических изолятов микобактерий с помощью теплового шока [18] или матрицы InstaGene (Bio-Rad Laboratories, Hercules, CA, USA). Анализ генов rpoB , secA1 , sodA и hsp65 был выполнен для идентификации на уровне подвидов, как описано ранее [12,13].Продукты ПЦР отправляли в Macrogen для секвенирования (Meibergdeef, 1105AZ, Амстердам, Нидерланды). Полученные последовательности анализировали с использованием алгоритма BLAST (NCBI).
Оценка генетической основы устойчивости
Гены erm (41) и rrl были проанализированы на устойчивость к кларитромицину. Ген erm (41) амплифицировали с использованием праймеров ermF (5’-GACCGGGGCCTTCTTCGTGAT-3 ’) и ermR (5’-GACTTCCCCGCACCGATTCC-3’) [7,8]. Ген rrl амплифицировали с использованием праймеров 19F (5’-GTAGCGAAATTCCTTGTCGG-3 ’) и 21R (5’-TTCCCGCTTAGATGCTTTCAG-3’) [7].Для гена erm (41) мы проанализировали полную последовательность гена из 673 п.н. и полиморфизм T28. Для гена rrl мы проанализировали фрагмент 836 п.н., включающий нуклеотиды 2058–2059, мутации которых вызывают устойчивость к кларитромицину.
Для амплификации erm (41) и rrl условия ПЦР составляли 5 минут при 95 ° C, затем 35 циклов при 95 ° C в течение 60 с, 62 ° C (для erm (41) , но 55 ° C). C для rrl ) в течение 60 с и 72 ° C в течение 90 с, а затем 72 ° C в течение 10 минут при окончательном удлинении.
Эталонные последовательности
Контрольные последовательности, использованные для классификации подвидов, были M . абсцесс subsp. абсцесс ATCC 19977T, M . абсцесс subsp. bolletii эталонный штамм CCUG 50184 (номера доступа GenBank AY859692 для rpoB и FJ442839.1 для secA1 ), M . абсцесс subsp. bolletii эталонный штамм CIP 108541 (инвентарные номера GenBank EU266576.1 для hsp65 и AY862403 для sodA ) и Mycobacterium abscessus subsp. massiliense CCUG 48898 (инвентарные номера GenBank AY593981 для rpoB , AY596465.1 для hsp65 , NZ_AKVF01000005.1 для secA1 и AY593975 для sodA ).
Эталонной последовательностью, использованной для анализа генов erm (41), и rrl , была M . абсцесс subsp. abscessus ATCC 19977T (инвентарный номер GenBank NC_010397).
Определение чувствительности к кларитромицину
Определение МПК кларитромицинапроводили в 9/16 исходных изолятах методом E-test (bioMérieux, bioMérieux España S.A., Мадрид, Испания) в чашках с агаром Мюллера-Хинтона при 30 ° C [19]. Тест на чувствительность к кларитромицину также был проведен в среде Мюллера-Хинтона с скорректированными катионами с использованием метода микроразбавления бульона [5] в чашках Sensititre RAPMYCOI (Sensititre, Trek Diagnostic Systems, East Grinstead, United Kindom) в соответствии с рекомендациями CLSI [9].Штаммы инкубировали в течение 14 дней, и МИК кларитромицина считывали через 3, 7 и 14 дней. Пороговое значение МПК кларитромицина для микроразведений колеблется от ≤2 чувствительности до ≥8 резистентности и ≤1 чувствительности до ≥4 резистентности для Е-теста [19].
Наблюдение за пациентом
Типирование штамма при последующем наблюдении за пациентом выполнялось с помощью анализа тандемных повторов (VNTR) с переменным числом. Используемые праймеры: TR45, TR109, TR116, TR150, TR155 и TR172. Шесть пар праймеров и локусов были такими же, как использовались Wong et al .[20]. Условия ПЦР для VNTR были следующими: начальная стадия денатурализации 15 мин при 95 ° C; 30 циклов денатурализации при 95 ° C в течение 60 с, отжига при 59 ° C в течение 60 с и удлинения при 72 ° C в течение 90 с, с заключительной стадией удлинения при 72 ° C в течение 10 мин. ПЦР-ампликоны анализировали с помощью системы капиллярного электрофореза EPOCH (BioTek, Германия). Результаты анализировали с помощью GeneMapper (Applied Biosystems).
Номера доступа последовательностей
Полученные последовательности для каждого гена были отправлены в GenBank и им были присвоены следующие номера доступа: от KT185536 до KT185557 для rpoB , от KT185514 до KT185535 для hsp65 , KT185603 до KT185624 для sodA , KT1856081 sec для sodA , KT18216081 sec для KT185492 — KT185513 для erm41 и KT185558 — KT185580 для rrl .
Результаты
Классификация подвидов на основе множественных генов
Анализ исходного изолята от каждого пациента выявил 11/16 M . абсцесс subsp. абсцесс (68,8%), 4/16 М . абсцесс subsp. боллетий (25,0%) и 1/16 М . абсцесс subsp. massiliense (6,3%). Для всех изолятов, кроме одного, мы наблюдали согласие между последовательностями генов rpoB , secA1 , sodA и hsp65 .Один штамм (изолят 08I004 из таблицы 1) был M . абсцесс subsp. abscessus согласно анализу rpoB , sodA и hsp65 , но M . абсцесс subsp. bolletii по гену secA1 . В таблице 1 показана подвидовая классификация 16 первых клинических изолятов, включенных в исследование.
Генетические основы устойчивости к кларитромицину
М . абсцесс комплексТаблица 1 также показывает результаты rrl и erm (41) и тип устойчивости исходных изолятов.Ни в одном из этих изолятов не наблюдали мутаций в гене rrl . Для полиморфизма erm (41) T28 мы идентифицировали 8 M . абсцесс subsp. абсцесс и 3 М . абсцесс subsp. боллетий . Для полиморфизма erm (41) C28 мы идентифицировали 3 M . абсцесс subsp. абсцесс . Мы обнаружили усеченный ген erm (41) в 1 M . абсцесс subsp. massiliense и 1 M . абсцесс subsp. bolletii , в каждом случае с двумя делециями в положениях 64 и 65 и окончательной делецией 276 п.н., как описано в литературе для M . абсцесс subsp. massiliense [16].
Впервые, насколько нам известно, в этом исследовании был обнаружен ген erm (41) , усеченный в 1 M . abscessus subsp bolletii .Данные о сходстве последовательностей генов rpoB , secA1 , sodA и hsp65 этого штамма и эталонных штаммов М . abscessus комплекс представлен в таблице 2.
Определение чувствительности к кларитромицину
Мы извлекли 9 штаммов M . абсцесс группа для фенотипического анализа. Методы Е-теста и микроразбавления были выполнены с восстановленными штаммами, чтобы сравнить эти два метода и связать результаты in vitro МИК с нашим генотипическим анализом.В таблице 1 приведены МИК кларитромицина для Е-теста и микроразбавления. Все штаммы были чувствительны к кларитромицину при использовании обоих методов через 3 дня. Пять из девяти штаммов были устойчивы к обоим методам через 7 дней, а два штамма были чувствительными. Один штамм был устойчив к Е-тесту и восприимчив к микроразведению, оставшийся штамм был восприимчив к Е-тесту и устойчив к микроразбавлению (Таблица 1). На 14-й день 7/9 штаммов оказались устойчивыми при использовании обоих методов. В обоих методах чувствительные и устойчивые штаммы согласовывались на 14-й день.
Наблюдение за пациентом
Таблица 1 также описывает результаты наблюдения за 4 пациентами: M . абсцесс subsp. abscessus штаммов, полученных от 3 пациентов (пациенты 1-3), показали приобретение точечной мутации в гене rrl . Пациент 1, erm (41) C28 sequevar, показал приобретенную устойчивость в позиции A2058G. Пациент 3, erm (41) T28 sequevar, показал приобретенную резистентность в позиции A2059G. Секевар erm (41) T28 от пациента 2 показал двойной пик в положении A2057G / A в последовательности ДНК, что указывает на новую выбранную мутацию конститутивной устойчивости к кларитромицину.В отличие от M . абсцесс subsp. Штамм bolletii от пациента 4 не обнаружил приобретенной мутации в гене rrl .
Все штаммы, изученные в ходе наблюдения, были идентичны исходным изолятам для каждого пациента. У каждого пациента был свой штамм. В таблице 1 также показан профиль переменного числа тандемных повторов анализа.
Обсуждение
Кларитромицин является ключевым агентом в лечении M . абсцесс комплекс инфекций [9]. Основная причина неэффективности лечения — индуцируемая резистентность [8,21]. Существенные различия в гене erm (41) , ассоциированном с индуцибельной устойчивостью, обнаружены среди M . abscessus комплекс [15,22], предполагая, что точная идентификация аллеля erm (41 ) важна для прогнозирования результатов лечения. Как и тройка М . Подвиды комплекса abscessus тесно связаны и демонстрируют горизонтальный перенос генов [11,23], для идентификации подвидов требуется более одного гена домашнего хозяйства [13].Тем не менее, используемые гены не определены [24]. Цель данной работы — охарактеризовать молекулярные механизмы устойчивости к кларитромицину у M . abscessus и проверить взаимосвязь между этими механизмами и тестом на чувствительность к кларитромицину.
В отличие от некоторых авторов [12,13] и в согласии с Nie et al. . [25], мы обнаружили, что каждый ген ( rpoB , secA1 , sodA и hsp65 ) мог классифицировать M . abscessus подвид, за исключением одного штамма, который был идентифицирован как M . абсцесс subsp. abscessus по rpoB , sodA и hsp65 и M . абсцесс subsp. bolletii по secA1 ген. В отличие от наших результатов, Tan et al . [26] смогли правильно классифицировать все свои штаммы, секвенировав ген secA1 . М . абсцесс subsp. abscessus был наиболее преобладающим подвидом комплекса (68,8%), в соответствии с данными литературы [12,21,27], где он колеблется от 51,2% до 78,5%. Следующим по преобладанию был М . абсцесс subsp. bolletii (25,0%), в отличие от многих работ, в которых вторым по частоте встречаемости подвидом является M . абсцесс subsp. massiliense [16,28].
Что касается механизмов генетической устойчивости, то секвенирование гена rrl в исходных изолятах показало отсутствие мутаций.Это контрастирует с предыдущими сообщениями [21,29], где наличие конститутивной резистентности к кларитромицину колеблется от 2,7% до 28,6%. Однако в соответствии с литературными данными мы наблюдали индуцибельную резистентность, обусловленную нуклеотидом T28 erm (41) в 68,8% (11/16) M . abscessus комплекс изолятов [8,21].
Как сообщалось ранее, erm (41) является специфическим для подвида геном в M . абсцесс группа [7,16]. Мы обнаружили только одно несоответствие опубликованным критериям [7,17], относящееся к M . абсцесс subsp. bolletii с укороченным геном erm (41) . Насколько нам известно, это первое описание M . абсцесс subsp. bolletii с нефункциональным усеченным геном erm (41) . Штамм (07I004) соответствовал M . абсцесс subsp. bolletii эталонных штаммов CCUG 50184 и CIP108541 для четырех изученных генов домашнего хозяйства, но не для гена erm (41) .Это открытие подтверждает другие сообщения, предполагающие, что перенос генов и рекомбинация происходят между подвидами [11,28]. Несколько корпусов М . абсцесс subsp. Сообщалось о massiliense с функциональным геном erm (41) [30]. Таким образом, наши данные подтверждают мнение о том, что erm (41) не является геном, специфичным для подвида, и что секвенирование гена erm (41) не должно использоваться в качестве единственного метода для классификации M . абсцесс сложный подвид.
Данные, полученные в результате нашего исследования, выявили приобретение конститутивной резистентности к кларитромицину. Конститутивная устойчивость к кларитромицину из-за мутаций в rrl была выбрана в штаммах erm (41) C28 (как показано у пациента 1) [7] и erm (41) T28 (как показано у пациентов 2 и 3). [21]. Благодаря преимуществу наличия исходных штаммов rrl дикого типа, наши результаты подтверждают эту идею, в отличие от Bastian et al . [7], отбор мутантов rrl аналогичен у штаммов T28 и C28.
Мы описали приобретение точечной мутации 2057 в T28 M . абсцесс subsp. abscessus с исходным изолятом rrl дикого типа. Наблюдался двойной пик в последовательности ДНК 2057 г., что предполагает сосуществование двух смешанных популяций ДНК ( rrl дикого типа и 2057 мутировавшая популяция), поскольку эти микобактерии обладают только одной копией гена rrl в организме человека. их геном [5]. Амплификацию и секвенирование ДНК повторяли три раза независимо, чтобы исключить ошибки амплификации или секвенирования.Дальнейшая работа должна включать клонирование, чтобы определить, существуют ли два варианта в этом штамме.
Важно подчеркнуть, что мутации в положении 2057 rrl у других микроорганизмов связаны со средне-низким уровнем устойчивости к кларитромицину in vitro [31], как видно из этого исследования. Вестер и др. . [31] также сообщили, что мутации в положениях 2057 и 2611 гена rrl вызывают устойчивость к макролидам и кетолидам из-за их близости к центру действия.Мутации в этих положениях препятствуют связи между антибиотиком и его мишенью и вызывают низкий уровень устойчивости к макролидам с 14-членным кольцом и отсутствие устойчивости к макролидам с 16-членным кольцом. Напротив, недавно Луо и др. . [32] сообщили о 2057 точечных мутациях в M . абсцесс subsp. abscessus , который показал высокий уровень устойчивости (MIC> 256 мкг / мл).
В нашем исследовании мы обнаружили полное согласие между секвенированием erm (41) , rrl и тестированием чувствительности к кларитромицину.После 3-дневного культивирования все штаммы были чувствительны, как это наблюдалось с помощью обоих методов (Е-тест и микроразведение). После 7-дневного культивирования было обнаружено 85% (6/7) индуцибельной устойчивости [ erm (41) T28 sequevar], а после 14-дневного культивирования мы обнаружили все индуцибельные штаммы устойчивости. Хотя мы не обнаружили существенных различий между этими двумя методами, у одного штамма с Е-тестом и у одного штамма с методом микроразбавления потребовалось 14 дней, чтобы выявить устойчивость. Это контрастирует с данными других авторов, которые сообщают о различиях между показателями Е-теста и микроразведения [33].Хотя микроразведение является стандартным методом в соответствии с CLSI [9], в нашем исследовании Е-тест показал те же результаты, что и микроразведение, и определение МИК на ранних стадиях было легче.
Мы хотели бы подчеркнуть, что одним из ограничений нашего исследования является относительно небольшое количество включенных изолятов. Это важно учитывать, делая выводы по эпидемиологии, но это не отменяет молекулярных наблюдений.
Заключение
В заключение, наши результаты выявили один штамм M . абсцесс subsp. bolletii с усеченным и нефункциональным erm (41) геном. Следует соблюдать осторожность, если секвенирование гена erm (41) — единственный метод, используемый для идентификации подвидов. Хотя конститутивная резистентность к кларитромицину не выявляется у исходных изолятов, она может быть приобретена у штаммов T28 и C28 erm (41) из M . абсцесс сложный. Наконец, приобретение мутации в положении 2057 в гене rrl придает устойчивость к кларитромицину от среднего до низкого уровня.
Благодарности
Авторы благодарят доктора Julià Gonzalez (Hospital Clinic Barcelona) за его поддержку метода микроразведения и Кэролайн Ньюи за руководство написанием рукописи.
Вклад авторов
Задумал и спроектировал эксперименты: ПК. Проведены эксперименты: MR FM MG CM ME. Проанализированы данные: MR FM PC. Предоставленные реагенты / материалы / инструменты анализа: MR FM MG CM ME PC. Написал статью: MR PC.
Ссылки
- 1.Гриффит Д.Е., Жирар В.М., Уоллес Р.Дж. Клинические особенности заболеваний легких, вызванных быстрорастущими микобактериями. Анализ 154 больных. Am Rev Respir Dis. 1993; 147: 1271–8. pmid: 8484642
- 2. Уоллес Р.Дж., Свенсон Дж. М., Силкокс В. А., Хороший RC, Чен Дж. А., Стоун МС. Спектр болезней, вызванных быстрорастущими микобактериями. Rev Infect Dis. 5: 657–79. pmid: 6353528
- 3. Нессар Р., Камбау Э., Рейрат Дж. М., Мюррей А., Гикель Б. Mycobacterium abscessus : новый кошмар антибиотиков.J Antimicrob Chemother. 2012; 67: 810–8. pmid: 222
- 4. Ли Ш., Ю ХК, Ким Ш., Ко В-Дж, Ким С.К., Пак Й.К. и др. Профиль лекарственной устойчивости штаммов группы Mycobacterium abscessus из Кореи. Ann Lab Med. 2014; 34: 31–7. pmid: 24422193
- 5. Гриффит Д.Е., Аксамит Т., Браун-Эллиотт Б.А., Катандзаро А., Дейли С., Гордин Ф. и др. Официальное заявление ATS / IDSA: диагностика, лечение и профилактика нетуберкулезных микобактериальных заболеваний.Am J Respir Crit Care Med. 2007. 175: 367–416. pmid: 17277290
- 6. Браун-Эллиот Б.А., Нэш К.А., Уоллес Р.Дж. Тестирование чувствительности к противомикробным препаратам, механизмы устойчивости к лекарствам и терапия инфекций нетуберкулезными микобактериями. Clin Microbiol Rev.2012; 25: 545–582. pmid: 22763637
- 7. Bastian S, Veziris N, Roux A-L, Brossier F, Gaillard J-L, Jarlier V и др. Оценка чувствительности к кларитромицину у штаммов, принадлежащих к группе Mycobacterium abscessus , с помощью секвенирования erm (41 ) и rrl .Антимикробные агенты Chemother. 2011; 55: 775–81. pmid: 21135185
- 8.
Нэш К. А., Браун-Эллиот Б. А., Уоллес Р. Дж. Новый ген, erm (41) , придает индуцибельную устойчивость к макролидам клиническим изолятам Mycobacterium abscessus , но отсутствует в Mycobacterium chelonae . Антимикробные агенты Chemother. 2009; 53: 1367–76. pmid: 1
99
- 9. Институт клинических и лабораторных стандартов, 2011. M24-A2: Тестирование чувствительности микобактерий, нокардий и других аэробных актиномицетов; Утвержденное стандартное второе издание — M24A2.
- 10. Леао СК, Тортоли Э., Эузеби ДжП, Гарсия МДж. Предложение о том, чтобы Mycobacterium massiliense и Mycobacterium bolletii были объединены и реклассифицированы как Mycobacterium abscessus subsp. боллетий гребень . ноя ., Обозначение Mycobacterium abscessus subsp. абсцесс subsp. ноя и исправленное описание Mycobacteri. Int J Syst Evol Microbiol. 2011; 61: 2311–3. pmid: 21037035
- 11.Сасси М., Дранкур М. Анализ генома выявляет три геномовида в Mycobacterium abscessus . BMC Genomics. 2014; 15: 359. pmid: 24886480
- 12. Желязны А.М., Рут Дж. М., Ши Ю. Р., Коломбо Р. Э., Шампута И. К., Сток Ф. и др. Когортное исследование молекулярной идентификации и типирования Mycobacterium abscessus , Mycobacterium massiliense , и Mycobacterium bolletii . J Clin Microbiol. 2009; 47: 1985–95. pmid: 19420162
- 13.Macheras E, Roux A-L, Ripoll F, Sivadon-Tardy V, Gutierrez C, Gaillard J-L, et al. Неточность секвенирования одной мишени для различения видов группы Mycobacterium abscessus . J Clin Microbiol. 2009. 47: 2596–600. pmid: 19515839
- 14. Fangous M-S, Mougari F, Gouriou S, Calvez E, Raskine L, Cambau E, et al. Алгоритм классификации для идентификации подвидов в пределах вида Mycobacterium abscessus , основанный на матричной лазерной десорбции / ионизационной времяпролетной масс-спектрометрии.J Clin Microbiol. 2014; pmid: 25009048
- 15. Браун-Эллиотт Б.А., Васиредди С., Васиредди Р., Яхьяева Э., Ховард С.Т., Нэш К. и др. Полезность секвенирования гена erm (41) в изолятах Mycobacterium abscessus subsp. abscessus с низкими и промежуточными МПК кларитромицина. J Clin Microbiol. 2015; 53: 1211–5. pmid: 25653399
- 16. Ким Х-И, Ким БДЖБ-Дж, Кук И-ХЙ, Юн И-Дж, Шин Дж. Mycobacterium massiliense отличается от Mycobacterium abscessus и Mycobacterium bolletii по гену (erm) метилтрансферазы эритромицин-рибосомы и типам чувствительности к кларитромицину.Microbiol Immunol. 2010; 54: 347–53. pmid: 20536733
- 17. Ёсида С., Цуюгути К., Сузуки К., Томита М., Окада М., Шимада Р. и др. Быстрая идентификация штаммов, принадлежащих к группе Mycobacterium abscessus , с помощью пиросеквенирования гена erm (41) . Диагностика Microbiol Infect Dis. Elsevier Inc .; 2014; 1–6.
- 18. Zhang ZQ, Ishaque M. Оценка методов выделения ДНК из медленно и быстро растущих микобактерий. Int J Lepr Other Mycobact Dis.1997: 65: 469–76. pmid: 9465157
- 19. Биле ЙОНР, Cavalieri SJ, Saubolle MA. Оценка Etest для тестирования чувствительности быстрорастущих микобактерий. К ним относятся: Оценка Etest для тестирования чувствительности быстрорастущих микобактерий. 1995; 33.
- 20. Вонг Ю.Л., Онг К.С., Нгеу Ю.Ф. Молекулярное типирование Mycobacterium abscessus на основе полиморфизма тандемных повторов. J Clin Microbiol. 2012; 50: 3084–8. pmid: 22760048
- 21.Ли С.Х., Ю Х.К., Ким С.Х., Кох В-Дж, Ким С.К., Пак Ю.К. и др. Обнаружение и оценка индуцируемых кларитромицином устойчивых штаммов среди корейских Mycobacterium abscessus Клинические штаммы: методы ПЦР. Анал J Clin Lab. 2014; 6: 1–6.
- 22. Ко WJ, Jeon K, Lee NY, Kim BJ, Kook YH, Lee SH и др. Клиническое значение дифференциации Mycobacterium massiliense от Mycobacterium abscessus . Am J Respir Crit Care Med. 2011; 183: 405–10.pmid: 20833823
- 23. Choo SW, Wee WY, Ngeow YF, Mitchell W, Tan JL, Wong GJ и др. Геномная разведка клинических изолятов нового патогена человека Mycobacterium abscessus показывает высокий эволюционный потенциал. Sci Rep.2014; 4: 4061. pmid: 24515248
- 24. Blauwendraat C, Dixon GLJ, Hartley JC, Foweraker J, Harris K A. Использование метода секвенирования с двумя генами для точного различения видов в комплексе Mycobacterium abscessus и Mycobacterium chelonae .Eur J Clin Microbiol Infect Dis. 2012; 31: 1847–53. pmid: 22222989
- 25. Nie W., Duan H, Huang H, Lu Y, Bi D, Chu N. Идентификация видов с использованием rpoB и hsp65 и тестирование чувствительности к восьми антибиотикам из Mycobacterium abscessus subsp. abscessus и Mycobacterium abscessus subsp. боллетий . Int J Infect Dis. 2014; pmid: 24932856
- 26. Тан JL, Khang TF, Ngeow YF, Choo SW.Филогеномный подход к классификации бактериальных подвидов: подтверждение концепции в Mycobacterium abscessus . BMC Genomics. 2013; 14: 879. pmid: 24330254
- 27. Ким Х-И, Кук И, Юн И-Дж, Пак К. Г., Ли Нью-Йорк, Шим Т.С. и др. Доли штаммов Mycobacterium massiliense и Mycobacterium bolletii среди корейских изолятов группы Mycobacterium chelonae-Mycobacterium abscessus . J Clin Microbiol. 2008; 46: 3384–90. pmid: 18753344
- 28.Чо Й-Дж, Йи Х, Чун Дж, Чо С-Н, Дейли С.Л., Ко В-Дж и др. Геномная последовательность штамма « Mycobacterium massiliense » CIP 108297 предполагает независимый таксономический статус комплекса Mycobacterium abscessus на уровне подвидов. PLoS One. 2013; 8: e81560. pmid: 24312320
- 29. Ёсида С., Цуюгути К., Сузуки К., Томита М., Окада М., Хаяси С. и др. Дальнейшее выделение Mycobacterium abscessus subsp. abscessus и subsp. bolletii в разных регионах Японии и чувствительность этих изолятов к антимикробным препаратам. Int J Antimicrob Agents. Elsevier B.V .; 2013; 42: 226–31.
- 30. Грей Т.Дж., Конг Ф., Джелфс П., Синченко В., Чен С.К. Улучшенная идентификация быстрорастущих микобактерий с помощью ПЦР с внутренней транскрибированной спейсерной областью 16S-23S и электрофореза в капиллярном геле. PLoS One. 2014; 9: e102290. pmid: 25013955
- 31. Вестер Б., Даутвейт С. Устойчивость к макролидам, обусловленная заменами оснований в 23S рРНК MINIREVIEW Antimicrob Agents Chemother.2001; 45: 1–12.
- 32. Луо Р.Ф., Карри С., Тейлор Н., Будвитене И., Банаи Н. Быстрое определение приобретенной и индуцируемой устойчивости к кларитромицину в группе Mycobacterium abscessus с помощью простого ПЦР-анализа в реальном времени. J Clin Microbiol. 2015; 1–12. pmid: 252
- 33. Woods GL, Bergmann JS, Witebsky FG, Fahle GA, Boulet B, Plaunt M и др. Многосайтовая воспроизводимость Etest для тестирования чувствительности Mycobacterium abscessus , Mycobacterium chelonae и Mycobacterium fortuitum .J Clin Microbiol. 2000. 38: 656–61. pmid: 10655363
За пределами промышленного комплекса тюрьмы Конференция 1998 г. — Критическое сопротивление
Название конференции:Критическое сопротивление: за пределами промышленного комплекса тюрьмы
Даты:25-27 сентября 1998 г.
Расположение:1 Калифорнийский университет в БерклиКритическое сопротивление было сформировано в 1997 году, когда активисты, оспаривающие идею о том, что тюремное заключение и полицейская деятельность являются решением социальных, политических и экономических проблем, собрались вместе, чтобы организовать конференцию, которая изучила и оспорила то, что мы стали называть тюрьмой. промышленный комплекс (ПИК).Первая конференция CR была способом начать кампанию (Stop Delano II, кампанию, которая остановила 20-летний бум строительства тюрем в Калифорнии), которая началась немедленно.
Запланированная бывшими заключенными, членами семьи, организаторами, учеными, адвокатами, поставщиками услуг, студентами и другими, цель конференции состояла в том, чтобы обратить внимание на тревожный рост тюремной системы, популяризировать идею «тюремно-промышленного комплекса». »(ПОС) и превратить« отмену »в практическую теорию изменений.
Конференция, проходившая в Беркли, Калифорния, в сентябре 1998 года, собрала более 3500 активистов, ученых, бывших и нынешних заключенных, лидеров профсоюзов, религиозных организаций, феминисток, активистов геев, лесбиянок и транссексуалов, молодежи, семей и политиков. буквально из любого штата и других стран. В трехдневном мероприятии приняли участие около 200 различных панелей и семинаров. Конференция также включала ряд культурных мероприятий и кинофестиваль.