


«Подольские курсанты» получили награду на фестивале военного кино в Туле
14 ноября в Тульском областном центре молодежи состоялась церемония закрытия XVIII Международного фестиваля военного кино имени Ю.Н. Озерова.
В этом году мероприятие прошло в онлайн-формате.
В номинации «Лучший фильм» победила картина «Подольские курсанты». Заместитель председателя правительства Тульской области Ольга Гремякова вручила «Золотой меч» сопродюсеру фильма Евгению Айзиковичу.
Также фильм получил награды в номинациях «Визуальные эффекты», «За лучший актерский ансамбль». Обладателем приза за лучшую мужскую роль стал Алексей Бардуков за роль лейтенанта Алёшкина фильме «Подольские курсанты».
Среди подольских курсантов были уроженцы Тульской области. По поручению губернатора Алексея Дюмина целый год учреждения культуры и Государственный архив Тульской области совместно проводили исследования и нашли сведения о 106 подольских курсантах — туляках. В 1941-м именно они были на Ильинском рубеже и защищали Москву, и большая часть из них погибла.
В этом году рядом с Тульским Суворовским училищем была открыта стела, на которой увековечены имена 106 подольских курсантов — туляков.
Приз за лучший фильм на иностранном языке вручила министр культуры Тульской области Татьяна Рыбкина. Победила картина «Восемь сыновей».
Министр отметила, что творческое сотрудничество между Тульской областью и Китайской Народной Республикой развивается. Проводятся совместные выставки, реализуются творческие проекты. Кроме того, планируется заключить договор о сотрудничестве с Нанкинским городским музеем и музеем «Тульский кремль».
В программе документального кино «Битва за жизнь» победила картина Владимира Сидоренко «Экспозиция войны».
Приз «За лучшую режиссуру» завоевал дебютант в игровом кино, режиссер фильма «Блокадный дневник» Андрей Зайцев.
Победитель в номинации «За правдивое отражение темы патриотизма в военном кино» — режиссер-постановщик фильма «Калашников» Константин Буслов.
Лучшими операторами жюри признало операторов-постановщиков фильма «Калашников» Левана Капанадзе и Максима Шинкоренко.
«Золотой меч» «За лучший сценарий» присужден автору фильма «Воины чести» Цинь Тянь. Приз получил представитель фильма, ведущий телеканала РЕН ТВ Игорь Прокопенко.
Приз Российского Союза ветеранов достался анимационному фильму тульского режиссера Дмитрия Чиркова «Сторожевой пёс Верный».
Фестиваль в Тульской области проходит уже в пятый раз. В этом году он прошел в дистанционном формате, а открытие и закрытие – как в онлайн-, так и в офлайн-формате. Показы шли на 23 площадках с учетом требований Роспотребнадзора – с заполняемостью зала не более 30%, сообщает пресс-служба правительства Тульской области.
В логове Белого Дракона / Хабр
В китайской мифологии «белый дракон» символизирует нечто непостижимое для человеческого воображения. Кадры в заголовке статьи до боли знакомы некоторым читателям. А тех, кому не особо узнаваемы — ждет удивительная экскурсия  Попытки создавать вечняк не только не сошли на нет в связи со стремительным развитием науки — наоборот их деятельность в последние годы набирает ход, привлекая всё новых и новых адептов. Что поделаешь, желание халявы и чудес в человеках неискоренимо. Такой размах движухи и послужил поводом к написанию этой статьи.
Попытки создавать вечняк не только не сошли на нет в связи со стремительным развитием науки — наоборот их деятельность в последние годы набирает ход, привлекая всё новых и новых адептов. Что поделаешь, желание халявы и чудес в человеках неискоренимо. Такой размах движухи и послужил поводом к написанию этой статьи.
Disclaimer: под катом дичь и надругательство над заблуждениями верующих
Известный всем со школы закон сохранения энергии в деле не помеха, так как справедлив исключительно для замкнутых систем. Но далеко не все системы являются изолированными как минимум от известных, а также еще не открытых наукой полей и излучений, пронизывающих пространство — что согласуется с научной точкой зрения, по крайней мере в качественном аспекте. И это обоснование является фундаментом искателей СЭ. Выходит что они по сути дела ищут
 Но по сложившейся традиции продолжают называть «вечняк», СЕ (сверхъединичник), а также БТГ (бестопливный генератор).
Но по сложившейся традиции продолжают называть «вечняк», СЕ (сверхъединичник), а также БТГ (бестопливный генератор).В одном только русскоязычном интернете исписаны многие тысячи страниц, в сотнях тем на множестве форумов. Некоторые ветки тянутся годами, но ничего по существу, чтоб прочитать и сделать — там не найти, от слова вообще. Опубликованные чертежи и схемы не дадут сверхъединицы. Сообщества иерархичны: как правило некий гуру ведет последователей шаг за шагом к постройке БТГ. Мотивы гуру не совсем понятны, возможно ЧСВ, самопиар, какие-то донаты. Но можно стать Гуру самостоятельно. Для этого необходимо:
- начальные навыки и опыт в электротехнике, своя домашняя лаборатория огромный плюс
-
свихнутьсяосвоить стиль общения, мышления, а также специфические термины в среде вечнякостоителей - твердо заявить на форумах о наличии у вас работающего сверхъединичника
Основные принципы гуру:
- уклоняться от принятой среди адекватно мыслящх людей конкретики, в любой её форме.
 Вопросы по существу, анализ, критика — считаются грубейшим моветоном. На сторонних ресурсах вне подконтрольных веток обсуждений необходимо быть предельно осторожным, там отсуствует возможность выпилить вопросы и мнения критиков.
Вопросы по существу, анализ, критика — считаются грубейшим моветоном. На сторонних ресурсах вне подконтрольных веток обсуждений необходимо быть предельно осторожным, там отсуствует возможность выпилить вопросы и мнения критиков. - строжайшая пошаговость: гуру предоставляет информацию семантически несвязанными порциями. Может давать задания не особо понятные для адептов — выполняя которые те получают шанс приблизиться к пониманию секретов СЭ. При этом планка понимания всегда отодвигается подобно горизонту.
- вечняк гуру не должен содержать локализованного сверхъединичного узла. Энергия там «разгоняется», циркулируя с самоусилением только по всему устройству в целом.
 Вопросы по существу, анализ, критика — считаются грубейшим моветоном. На сторонних ресурсах вне подконтрольных веток обсуждений необходимо быть предельно осторожным, там отсуствует возможность выпилить вопросы и мнения критиков.
Вопросы по существу, анализ, критика — считаются грубейшим моветоном. На сторонних ресурсах вне подконтрольных веток обсуждений необходимо быть предельно осторожным, там отсуствует возможность выпилить вопросы и мнения критиков.После чего вам обеспечен штат последователей, далее всё зависит исключительно от личной харизмы, навыков социальной инженерии, умения убедительно переобещивать обещанное. Одни гуру сливаются быстрее, другие умудряются водить толпу народа по пустыне на протяжении многих лет.
Среди искателей СЭ прослеживается тенденция: механик фанатеет по теории эфира, отличный на своем поприще слесарь пытается создать электрический вечняк, «нефизик» опирается на спорные физические принципы, а электрик обеспокоен рычагами и шестеренками.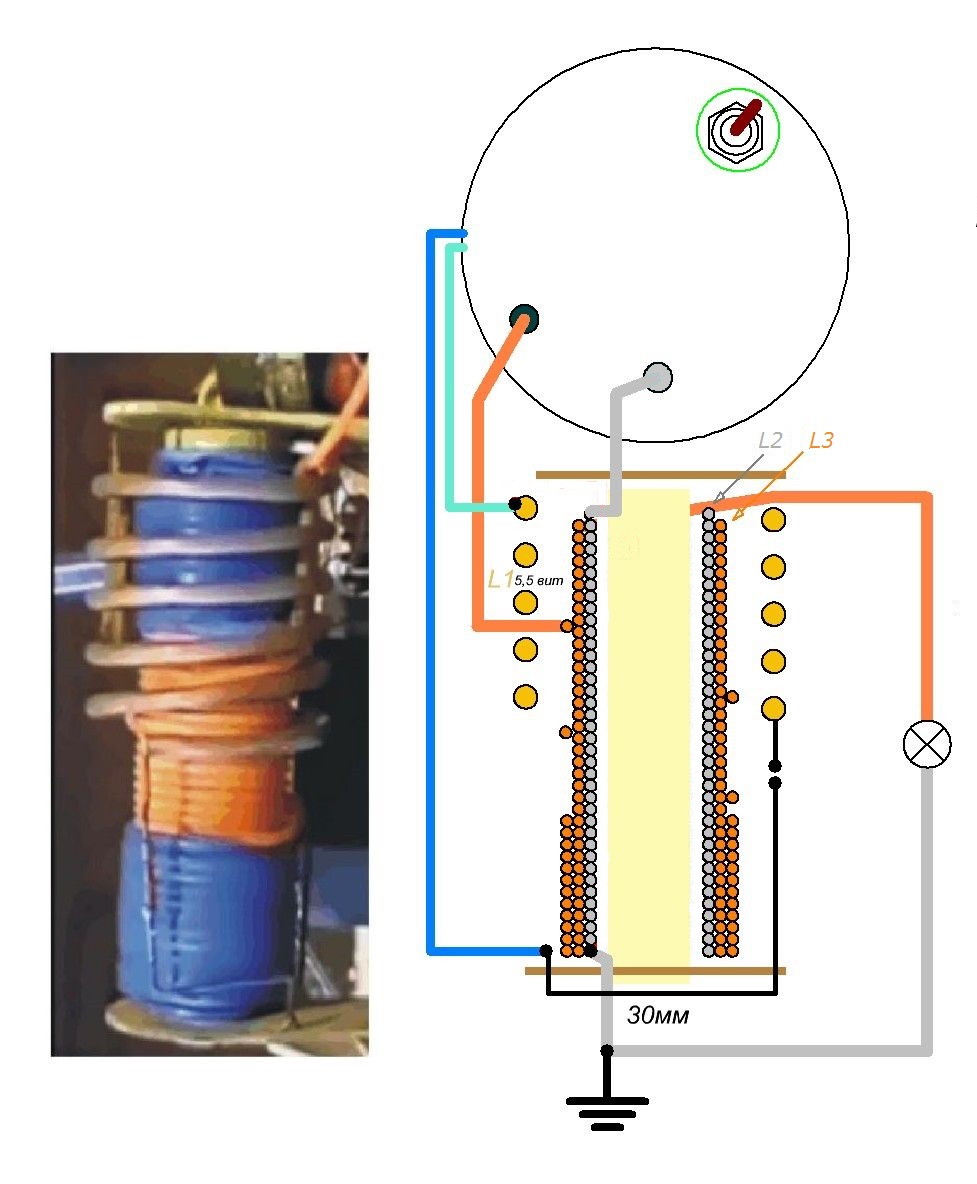
Второй аспект — пренебрежение научным методом. Можно еще понять веру в заговоры, порождающую недоверие к теориям современной науки и надежду на существование тщательно скрываемых запретных технологий получения энергии. Но методологию-то отвергать зачем? Простите судари — вы занимаетесь техническими исследованиями, а не вещанием с трибун. Железо не обманешь. В технике что-либо создать получится лишь объективно следуя научному подходу.
Далее примеры тематик поиска СЭ
Идеи и генераторы Николы Тесла
С позиции радиоинженера, в генераторе Тесла нет ничего особенного: обычный высоковольтный резонансный транформатор, работающий на частотах от сотен кГц до пары МГц. Однослойная высокодобротная катушка, являющаяся по совместительству объемным колебательным контуром, образованным индуктивностью и конструктивной межвитковой емкостью.
В основе всего — Рэзонансъ! © Капанадзе
Тариэль Капанадзе, демонстрируя свои технические фокусы на публике за небольшую плату плюс большой самопиар — вряд мог догадываться что невольно станет причиной наиболее элитного фундаменталистского течения среди российских вечнякостроителей. Как оказалось, именно его незамысловатая конструкция лучше всего подходит для становления гуру. Авторский БТГ Капанадзе не отличался особой изощренностью. Кое-как выпиленный ящик из дешевого пластика с бутафорской катушкой — банально питается от находящегося рядом автомобильного аккумулятора.
Но тем не менее, благодаря своей харизматичности и особенностям местного менталитета, Капанадзе уверенно внушает зрителям что аппарат сверхъединичный. Завоевал широкую известность и популярность в желтых СМИ.
В отличие от Капанадзе, гуру российского пошива напрочь избегают публичной демонстрации своих репликаций БТГ, ограничиваясь исключительно видеороликами на ютубе, которых там несметное количество.
Отличительная черта вечнякостроителей этого направления — отсуствие сколь-нибудь вменяемых публикаций в форме технических статей. Снимают только видео, в большинстве своем с ужасным светом и трясущейся камерой, которая показывает путаницу проводов, деталей и катушек — где толком невозможно ничего понять. Хотя изредка встречаются видеоролики и без трясучки. А также отошли от палева с аккумулятором, который теперь заменяют хорошо замаскированными проводами, подводящими питание. Декларируемая концепция: формирование условий для синхронизации «вторых», «третьих» и прочих резонансов, получение особого бестокового и беспроводного электричества — что с точки зрения специалиста сплошная дичь в попытках по-своему объяснить общеизвестные в радиотехнике явления.
Мотор Бедини
Это обширный класс устройств, где авторы стремятся получить сверхъединичность путем утилизации ЭДС самоиндукции катушек. В надежде что одна обмотка будет раскручивать колесо, а энергию будем снимать с нескольких других, самоиндукцию которых тоже употребим на пользу дела. Встречаются с вращающимися частями как на фото, так и чисто электротехнические версии не содержащие механики. Подход везде один и тот же — основан на непонимании что катушка не выдаст больше энергии чем в неё вкачали, и что нагруженная обмотка генератора сопротивляется вращению, в отличе от ненагруженной. Абсолютно беспреспективное направление, не выдерживающее проверки на самозапитку — которая является основным критерием сверхъединичности.
Правда Бедини таки выкрутился аналогично предыдущему персонажу — поставил стеллажи аккумуляторов, способных питать устройство неделями. Куча красиво сделанных моделей БТГ успешно стимулирует последователей на репликации (не будут же такое строить если б не работало — ага), а также спонсоров на донаты для продолжения исследований.
Магнитные моторы Perendev, Minato и другие
Попытки запрячь консервативные силы магнитного поля, в свое время стимулировали спрос на неодимовые магниты со стороны тысяч экспериментаторов по всему миру. Указанные в заголовке авторы подняли неплохие бабки. Правда один из них впоследствии отправился в места не столь отдаленные по искам недовольных спонсоров, так и не получивших обещанный мотор. На практике магнитный двигатель работает только если держать магнит статора в руках — из-за рефлективных неосознанных движений. Стоит его закрепить, и всё насмарку.
Тем не менее некоторая польза от возни с магнитами имеется — волей-неволей экспериментаторы начинают разбираться и понимать их свойства. Тема хоть и неперспективная, но все же не наносящая катастрофического вреда мировоззрению.
Механический маятник Велько Милковича
Завершить этот небольшой обзор хотелось бы на позитивной ноте, поэтому выбрал максимально нешизовое, а главное не утаиваемое автором техническое решение. Насколько удалось понять, автор считает что сумел запрячь центробежную силу, создаваемую качающимся маятником в нижней части его траектории. Используется параметрический маятник на коромысле, точка подвеса маятника немного ходит вверх-вниз.
Насколько удалось понять, автор считает что сумел запрячь центробежную силу, создаваемую качающимся маятником в нижней части его траектории. Используется параметрический маятник на коромысле, точка подвеса маятника немного ходит вверх-вниз.
На видео одна из репликаций «усилителя Милковича» для раскачивания водяной помпы небольшими усилиями при подталкивании маятника. В дискуссиях автор неоднократно пояснял, что руками дернуть эту помпу совершенно не хватит сил.
Тема механики для меня непрофильная, но думаю что там нету сверхъединичности, хотя автор считает иначе. Тем не менее эта рацуха вполне может найти применение в хозяйстве.
На самом деле перечисленные выше примеры сферы поисков «сверхъединицы» — лишь капля в море из числа всех существующих. Базисом фриковых исследований до сих пор является теория эфира, в умах здравомыслящих людей уже сто лет назад закономерно уступившая место ОТО, КМ и КТП — которые успешно подтверждаются многими экспериментами. Но экспериментаторы все равно тратят время, силы, деньги, осуществляя в гаражах большие но заведомо провальные проекты. Немного пообщавшись в этой сфере, уже прекрасно понял что состоявшихся «исследователей СЭ» вряд ли получится в чем убедить. Скорее бесполезно. Есть мнение что возня с техникой в любом случае даёт развитие — но лично я его не разделяю, т.к. отравленное аномально-шизическими представлениями мышление практически теряет способность к аналитике и применению разумного научного подхода. Но может быть кому-то из начинающих, еще не погрязших в псевдонаучной аномальщине — данный материал в чем-то поможет разобраться…
Немного пообщавшись в этой сфере, уже прекрасно понял что состоявшихся «исследователей СЭ» вряд ли получится в чем убедить. Скорее бесполезно. Есть мнение что возня с техникой в любом случае даёт развитие — но лично я его не разделяю, т.к. отравленное аномально-шизическими представлениями мышление практически теряет способность к аналитике и применению разумного научного подхода. Но может быть кому-то из начинающих, еще не погрязших в псевдонаучной аномальщине — данный материал в чем-то поможет разобраться…
Эпатажный Саакашвили вернулся в Грузию, а его отправили в тюрьму
Омбудсмен Грузии прибыла в тюрьму, где находится Саакашвили, чтобы ознакомиться с состоянием его здоровья. Адвокат арестованного жалуется на то, что не может выйти с ним на связь. У руставской тюрьмы в эти минуты ажиотаж. В Тбилиси, где в одной из квартир и задержали беглого экс-президента, один за другим – брифинги. МВД сообщает, что Саакашвили прибыл в Грузию еще четыре дня назад. Как будет реагировать Тбилиси на вызов своего посла в МИД Украины? Зеленский уже сказал, что обеспокоен. И кому выгодны такие камбеки Саакашвили?
Как будет реагировать Тбилиси на вызов своего посла в МИД Украины? Зеленский уже сказал, что обеспокоен. И кому выгодны такие камбеки Саакашвили?
Саакашвили доставили в руставскую тюрьму в пригороде Тбилиси. Бывшего президента ведут по коридору под конвоем, он же совсем не выглядит разочарованным, скорее, наоборот.
Такой исход явно не был неожиданностью для заочно осужденного в Грузии Саакашвили. На его странице в Facebook появился ролик, где он говорит о возможном аресте.
«Меня могут сейчас задержать в Тбилиси, но вы должны знать, что мы сражаемся до конца», – говорит Саакашвили. Он добавил, что «3 [октября] все вместе будем праздновать победу».
А началось все с видео, где Саакашвили говорит: » Вот это мой Батуми».
Узнаваемый образ, хоть голова скрыта капюшоном, а на лице маска, спущенная в момент записи. Фигурант четырех уголовных дел в Грузии, Михаил Саакашвили объявил, что якобы вернулся на родину.
«Я так скучал по этим местам! Сегодня день исполнения моей мечты! Я рискнул всем: своей жизнью, свободой, чтобы быть тут.
 А от вас прошу одного – прийти на выборы», – призвал Саакашвили.
А от вас прошу одного – прийти на выборы», – призвал Саакашвили.Видео мгновенно стало скандальным. Как объявленный в розыск, известный всем беглый экс-президент мог попасть в Грузию? Хотя возникли сомнения в подлинности ночного, довольно размытого видео. Весь день в МВД республики заявляли, что госграницу ни Грузии, ни Украины Саакашвили не пересекал. Пользователи соцсетей тем временем обсуждали другой ролик: как колонна легкой бронетехники направляется в Батуми. К вечеру премьер страны собрал срочный специальный брифинг.
«Я хочу сообщить нашему обществу, что третий президент Грузии, разыскиваемый Михаил Саакашвили, задержан. У наших правоохранительных органов заранее была информация о его передвижении, начиная из Украины, заканчивая Грузией. Этот процесс был под полным контролем», – проинформировал глава правительства республики Ираклий Гарибашвили.
И все же, как Саакашвили смог проникнуть через грузинскую госграницу? Накануне в соцсетях он выложил фотографию забронированного себе и матери авиабилета по маршруту Киев-Тбилиси, планировал прибыть вечером в субботу. Теперь понятно, что это был отвлекающий маневр. Но попасть в Батуми в пятницу морем, как пишет «Спутник Грузия», Саакашвили не мог, так как там уже два дня шторм, и суда не заходят в порт. Впрочем, позднее появилась информация, что, несмотря на непогоду, в порту все же приняли два грузовых судна: из Румынии и Украины. Но тогда как среди команды матросов, мотористов и боцмана не узнали Саакашвили? В правящей партии «Грузинская мечта» уверены: все это – очередной политический трюк эпатажного экс-президента.
Теперь понятно, что это был отвлекающий маневр. Но попасть в Батуми в пятницу морем, как пишет «Спутник Грузия», Саакашвили не мог, так как там уже два дня шторм, и суда не заходят в порт. Впрочем, позднее появилась информация, что, несмотря на непогоду, в порту все же приняли два грузовых судна: из Румынии и Украины. Но тогда как среди команды матросов, мотористов и боцмана не узнали Саакашвили? В правящей партии «Грузинская мечта» уверены: все это – очередной политический трюк эпатажного экс-президента.
«Он клоун. Я хотел бы, чтобы он был здесь. В этом случае все общество увидело бы, каким клоуном является Михаил Саакашвили. И этим окончательно завершилось бы то уродство, которое есть в грузинской политике, в том числе с точки зрения поляризации», – заявил председатель правящей в стране партии «Грузинская мечта – Демократическая Грузия» Ираклий Кобахидзе.
Но история, похоже, только начинается. И момент для возвращения был выбран расчетливо: в стране кипят политические страсти в преддверии завтрашних местных выборов депутатов и глав городов. Его возвращение и неминуемый арест переворачивают политическую ситуацию в Грузии прямо накануне выборов. Оппозиция уже резко ужесточает риторику.
Его возвращение и неминуемый арест переворачивают политическую ситуацию в Грузии прямо накануне выборов. Оппозиция уже резко ужесточает риторику.
«Если он вернулся, то это доказывает то, что у нас нет государства, нет структур, нет границ, нет МВД, нет службы безопасности, которая прослушивает священнослужителей, вас, мирных граждан. Если это так, значит, государство разрушено», – подчеркнул лидер партии «Лело» Мамука Хазарадзе.
Очевидно, вырастет и внешнее давление. В Грузию на выборы направлены сотни наблюдателей с Запада, где у Саакашвили немало покровителей. В том числе и поэтому он многократно и так уверенно призывал своих сторонников завтра выходить на улицы.
«Дестабилизация, скорее всего, будет. Найдутся недовольные результатами выборов, будет заявлено о том, что выборы были сфальсифицированы. Повторится ситуация, которая сложилась после последних парламентских выборов», – прогнозирует политолог Васо Капанадзе.

Политическая дестабилизация – стихия Саакашвили. Он смог удивить даже бывалых на Украине, когда штурмом брал польско-украинскую границу или буквально убегал по крышам от полицейских, после того как Порошенко решил выдворить его с позором, лишив украинского паспорта.
Со стороны Саакашвили, как всегда, было много эпатажа, громких заявлений и поступков, которые так ничем и не закончились. А символом восьмилетних политических гастролей стала знаменитая перепалка с бывшим главой украинского МВД.
В грузинском обществе отношение к Саакашвили крайне неоднозначное. Теперь же его появление и арест могут спровоцировать ситуацию с непредсказуемым исходом.
Фактор роста соединительной ткани (CTGF / CCN2) опосредует ангиогенный эффект S1P в эндотелиальных клетках микрососудов кожи человека
(A) HDMEC были трансдуцированы аденовирусами Go , CTGF, CT и NT CTGF при MOI10 в течение 24 часов; 25 мкг общих белков разделяли с помощью SDS-PAGE для MMP1, MMP9 и переносили на нитроцеллюлозную мембрану.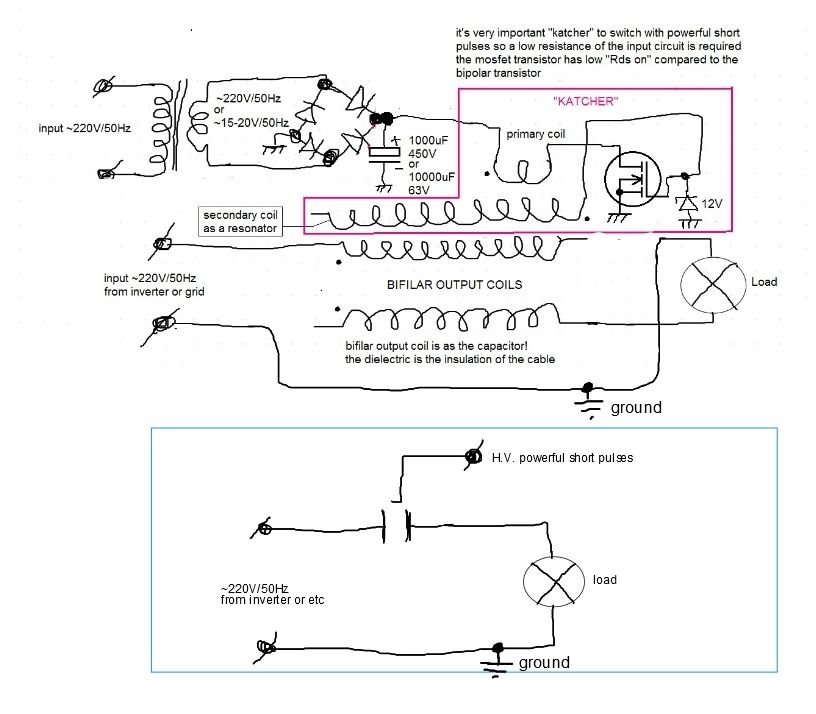 Блоты зондировали в течение ночи первичным Abs при 4 ° C. В качестве контроля для равной нагрузки белка мембраны были очищены и повторно исследованы на β-актин. (B) Графическое представление экспрессии белков MMP1 и MMP9, определенной с помощью вестерн-блоттинга, представленного в (A). Значения представляют собой среднее значение ± SEM трех независимых экспериментов, проведенных в двух экземплярах. * Значимые значения при p (C) HDMEC были трансдуцированы без сайленсинга (NS) или миРНК AdCTGF с добавлением S1P. Блокирование эндогенного CTGF ингибировало миграцию клеток, стимулированную S1P. (D) HDMEC трансдуцировали возрастающими дозами AdN-концевого (AdNT) и AdC-концевого (AdCT) домена CTGF, и уровень экспрессии определяли с помощью вестерн-блоттинга. (E) Графическое представление уровня экспрессии AdNT и AdCT представлено в (D). (F) HDMEC культивировали до 80% слияния в полной среде с последующей трансдукцией аденовирусами (AdGo, AdCTGF, AdNT и AdCT) или обрабатывали rhCTGF.
Блоты зондировали в течение ночи первичным Abs при 4 ° C. В качестве контроля для равной нагрузки белка мембраны были очищены и повторно исследованы на β-актин. (B) Графическое представление экспрессии белков MMP1 и MMP9, определенной с помощью вестерн-блоттинга, представленного в (A). Значения представляют собой среднее значение ± SEM трех независимых экспериментов, проведенных в двух экземплярах. * Значимые значения при p (C) HDMEC были трансдуцированы без сайленсинга (NS) или миРНК AdCTGF с добавлением S1P. Блокирование эндогенного CTGF ингибировало миграцию клеток, стимулированную S1P. (D) HDMEC трансдуцировали возрастающими дозами AdN-концевого (AdNT) и AdC-концевого (AdCT) домена CTGF, и уровень экспрессии определяли с помощью вестерн-блоттинга. (E) Графическое представление уровня экспрессии AdNT и AdCT представлено в (D). (F) HDMEC культивировали до 80% слияния в полной среде с последующей трансдукцией аденовирусами (AdGo, AdCTGF, AdNT и AdCT) или обрабатывали rhCTGF.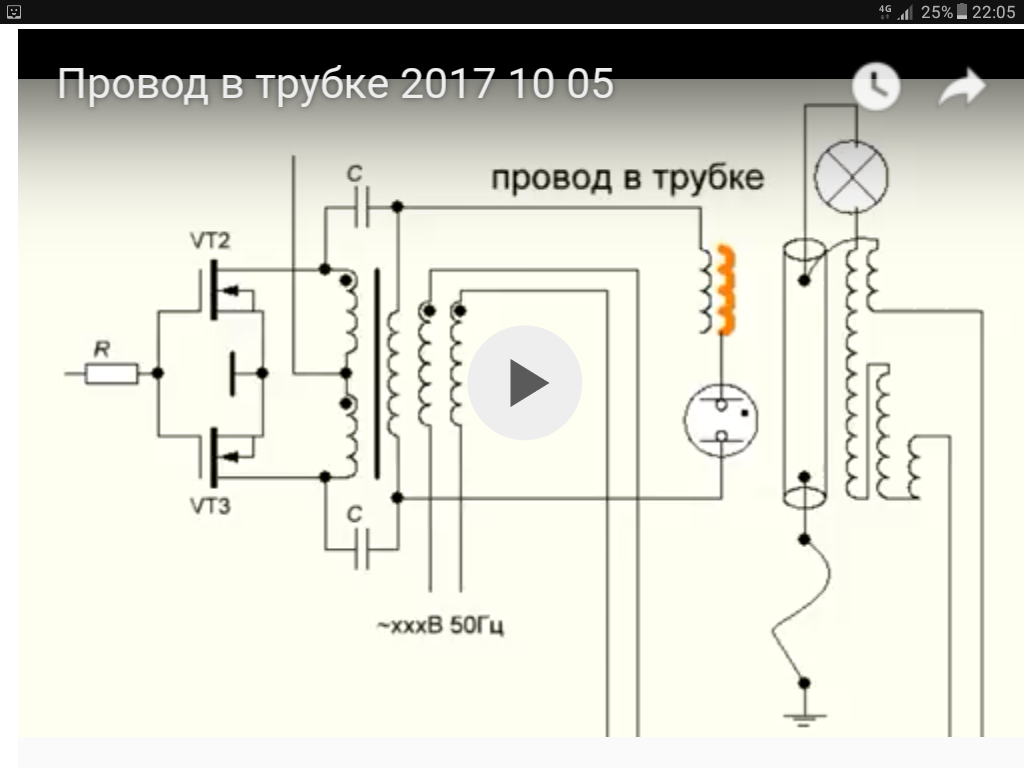 Через 8 часов клетки были механически «ранены» путем соскабливания наконечником марки Fisher. В течение следующих 24-72 часов клетки хранили в среде EBM2, содержащей 0,5% сыворотки. Представлены репрезентативные изображения. (G) Гистограмма представляет скорость миграции, выраженную как процент клеток, которые пересекли «царапину» по сравнению с идентичной «без царапиной» областью.Подсчет клеток производился через 48 часов после травмы. Эксперименты повторяли трижды в двух экземплярах. (H) Графическое представление анализа миграции при совместном культивировании через лунки. Вкратце, кожные фибробласты трансдуцировали указанными аденовирусами. На следующий день среду меняли и на вкладыши для культур клеток высевали HDMEC. Через 24 часа вставки удалили и подсчитали клетки, как описано в разделе «Методы». (I) Гистограмма показывает увеличение пролиферации эндотелиальных клеток после трансдукции клеток с помощью AdCTGF и AdCT.Значения представляют собой среднее значение ± SEM трех независимых экспериментов, проведенных в двух экземплярах.
Через 8 часов клетки были механически «ранены» путем соскабливания наконечником марки Fisher. В течение следующих 24-72 часов клетки хранили в среде EBM2, содержащей 0,5% сыворотки. Представлены репрезентативные изображения. (G) Гистограмма представляет скорость миграции, выраженную как процент клеток, которые пересекли «царапину» по сравнению с идентичной «без царапиной» областью.Подсчет клеток производился через 48 часов после травмы. Эксперименты повторяли трижды в двух экземплярах. (H) Графическое представление анализа миграции при совместном культивировании через лунки. Вкратце, кожные фибробласты трансдуцировали указанными аденовирусами. На следующий день среду меняли и на вкладыши для культур клеток высевали HDMEC. Через 24 часа вставки удалили и подсчитали клетки, как описано в разделе «Методы». (I) Гистограмма показывает увеличение пролиферации эндотелиальных клеток после трансдукции клеток с помощью AdCTGF и AdCT.Значения представляют собой среднее значение ± SEM трех независимых экспериментов, проведенных в двух экземплярах. * Значимые значения при p
* Значимые значения при p Рисунок 3. AdCTGF siRNA отменяет S1P-индуцированный спинной мозг…
Рисунок 3.AdCTGF siRNA отменяет S1P-индуцированное образование пуповины
(A) При 80% конфлюэнтности HDMEC были наложены…
Фигура 3. AdCTGF siRNA отменяет S1P-индуцированное образование пуповины. (A) При 80% конфлюэнтности HDMEC покрывали коллагеном и инкубировали в 0,5% EBM2 и, где указано, с добавлением S1P (500 нМ) или VEGF в течение 24-48 часов (эндотелиальные клетки метили липофильными живыми клетками). краситель CM Dil (1 мкг / мл) [молекулярные зонды]. В присутствии VEGF и S1P капиллярно-подобные тяжи образовывались через 24 часа, в то время как эндотелиальные клетки в SFM не образовывали тяжей. Опыты повторяли трижды. (B) Графическое представление длины шнура и количества ячеек представлено в ( A ). (C) HDMEC трансдуцировали с помощью немолчания (NS) или миРНК AdCTGF в течение 24 часов без добавления или с добавлением S1P. (D) Количественная оценка образования корда, наблюдаемого в (C), выполняется, как описано в разделе «Методы».Значения представляют собой среднее ± SEM трех независимых экспериментов, проведенных в двух экземплярах. * Значимые значения при p <0,05
В присутствии VEGF и S1P капиллярно-подобные тяжи образовывались через 24 часа, в то время как эндотелиальные клетки в SFM не образовывали тяжей. Опыты повторяли трижды. (B) Графическое представление длины шнура и количества ячеек представлено в ( A ). (C) HDMEC трансдуцировали с помощью немолчания (NS) или миРНК AdCTGF в течение 24 часов без добавления или с добавлением S1P. (D) Количественная оценка образования корда, наблюдаемого в (C), выполняется, как описано в разделе «Методы».Значения представляют собой среднее ± SEM трех независимых экспериментов, проведенных в двух экземплярах. * Значимые значения при p <0,05
Рисунок 3. AdCTGF siRNA отменяет S1P-индуцированный спинной мозг…
Фигура 3. AdCTGF siRNA отменяет S1P-индуцированное образование пуповины
AdCTGF siRNA отменяет S1P-индуцированное образование пуповины
(A) При 80% конфлюэнтности HDMEC были наложены…
Рисунок 3.AdCTGF siRNA отменяет S1P-индуцированное образование пуповины (A) При 80% конфлюэнтности HDMEC покрывали коллагеном и инкубировали в 0,5% EBM2 и, где указано, с добавлением S1P (500 нМ) или VEGF в течение 24-48 часов (эндотелиальные клетки метили липофильными живыми клетками). окраска CM Dil (1 мкг / мл) [Molecular Probes]. В присутствии VEGF и S1P капиллярно-подобные тяжи образовывались через 24 часа, в то время как эндотелиальные клетки в SFM не образовывали тяж. Эксперименты повторяли три раза. (B) Графическое представление длины шнура и количества ячеек представлено в ( A ). (C) HDMEC трансдуцировали с помощью немолчания (NS) или миРНК AdCTGF в течение 24 часов без добавления или с добавлением S1P. (D) Количественная оценка образования корда, наблюдаемого в (C), выполняется, как описано в разделе «Методы». Значения представляют собой среднее ± SEM трех независимых экспериментов, проведенных в двух экземплярах. * Значимые значения при p <0,05
(D) Количественная оценка образования корда, наблюдаемого в (C), выполняется, как описано в разделе «Методы». Значения представляют собой среднее ± SEM трех независимых экспериментов, проведенных в двух экземплярах. * Значимые значения при p <0,05
Рисунок 4.Повышенная экспрессия N-конца и C-конца…
Рисунок 4. Повышенная экспрессия CTGF на N-конце и C-конце индуцирует образование капиллярных тяжей
(A) HDMEC были…
Рисунок 4. Повышенная экспрессия CTGF на N-конце и C-конце индуцирует образование капиллярных тяжей. (A) HDMEC трансдуцировали с помощью AdCTGF, AdNT или AdCT или контрольного AdGo. На следующий день была создана совместная 3D-культура, как описано в разделе «Методы». Клетки инкубировали в 0,5% EBM2 в течение 24-48 часов. Опыты повторяли трижды. (B) Графическое изображение длины трубки и количества ячеек, представленных на (A). Значения представляют собой среднее ± SEM трех независимых экспериментов, проведенных в двух экземплярах. * Значимые значения при p <0,05
На следующий день была создана совместная 3D-культура, как описано в разделе «Методы». Клетки инкубировали в 0,5% EBM2 в течение 24-48 часов. Опыты повторяли трижды. (B) Графическое изображение длины трубки и количества ячеек, представленных на (A). Значения представляют собой среднее ± SEM трех независимых экспериментов, проведенных в двух экземплярах. * Значимые значения при p <0,05
Принцип работы Капанадзе
Если мы сможем понять мотивы, мы могли бы написать симфонию… Э. Дога Принцип работы аппарата Тариела Капанадзе
Просмотры 148 Загрузки 70 Размер файла 921KB
Отчет DMCA / Copyright
СКАЧАТЬ ФАЙЛ
Рекомендовать историиПредварительный просмотр цитирования
Если мы сможем понять мотивы, мы могли бы написать симфонию … E. Doga
Принцип работы устройства Тариэла Капанадзе
Первоначально опубликовано на русском языке: Виктор Григ — март 2009 г. Перевод: Стойчо Стоев (Дата: 23 января 2009 г.)
Перевод: Стойчо Стоев (Дата: 23 января 2009 г.)
1
I.
МОРАЛЬНЫЙ АСПЕКТ
После просмотра демонстрационного фильма Тариела Капанадзе о свободном извлечении энергии я начал испытывать симпатию к этому человеку как к преданному изобретателю, так и к человеку. Мы должны отдать должное ему уважение и поставить его на один уровень с Эдвином Греем, Джоном Бедини, Хаббардом, Б. Гребенниковым и остальными изобретателями бесплатных нервных исследований.
II.
СОЦИАЛЬНЫЙ АСПЕКТ
Это правда, что свободная энергия долго тянет за собой все последствия, на которые мы в настоящее время не вмешиваемся.Наибольшее влияние имеет то, что это устройство может заменить электростанцию или любой другой источник энергии. Следовательно, это может свести на нет все доходы от природных ресурсов в виде экспорта энергии и той части валового национального продукта, которая связана со всеми аспектами добычи и транспортировки целевого топлива. Жизнь на Земле постоянно развивается. У нас есть космические корабли, которые путешествуют по всей Солнечной системе, но здесь, на Земле, до сих пор все еще есть масса, использующая ископаемые топлива в качестве основного источника транспорта.Совершенно необходимо, чтобы мы, исследователи, принимали решения для собственной защиты. Вся информация об исследованиях свободной энергии должна будет поступать в широко открытый Интернет, чтобы она могла быть сохранена в низовых слоях свободного энергетического движения. III.
У нас есть космические корабли, которые путешествуют по всей Солнечной системе, но здесь, на Земле, до сих пор все еще есть масса, использующая ископаемые топлива в качестве основного источника транспорта.Совершенно необходимо, чтобы мы, исследователи, принимали решения для собственной защиты. Вся информация об исследованиях свободной энергии должна будет поступать в широко открытый Интернет, чтобы она могла быть сохранена в низовых слоях свободного энергетического движения. III.
ОСНОВНЫЕ МОТИВЫ
В своем объяснении я постараюсь полностью исключить присутствие воды, как показано в фильме. Принцип работы устройства Капанадзе основан на принципиальной взаимозависимости, и это основа всего устройства.Это довольно просто, но для объяснения нам придется построить некую концептуальную основу без каких-либо предубеждений.
2
На рисунке выше изображен патент Тариэла Капанадзе. Он выглядит совсем иначе, чем зеленый прямоугольник, который мы видим на видео, и, кроме того, похоже, что он был улучшен. IV.
IV.
ЕДИНЫЙ ЦИКЛ РЕЗОНАНСА В СРЕДЕ
Начнем с самого начала. Параллельный LC cerquit.
Зарядить конденсатор. Разрядите конденсатор в индукционную катушку.Индукция создает напряжение в непосредственной среде катушки — мягкий толчок, толчок, а не резкие импульсы. Среда толкает назад, вызывая самоиндукцию, разряжая конденсатор, вызывая, как следствие, резонанс 3
в цепи. Таким образом, цепь создает устойчивую волну, создающую резонансную частоту. В чем причина этого? Это следствие толчка окружающей среды в одну сторону и зарядки конденсатора за счет поглощения энергии, создаваемой последующим толчком окружающей среды в противоположную сторону.
Стопроцентный резонанс одного цикла на мосту, когда по нему маршируют римские солдаты. Серферс моста отталкивается, и этого достаточно, чтобы мост вошел в резонанс и сломался. Вернемся к сертификату LC. Я буду изменять емкость в возрастающем или убывающем направлении. Что происходит?
Резонанс существует постоянно, только его частота и амплитуда изменяются вследствие изменения «баланса энергии» в схеме. С точки зрения местной среды — все осталось, как было раньше. Он, как и прежде, имеет электромагнитное сопротивление, всегда пытаясь уравновесить созданный локальный градиент от напряжения, «упругости». В развитии этой мысли возникает вопрос. Есть ли естественный резонанс окружающей среды. В смысле, могли бы мы случайно, используя нашу электрическую схему, найти какой-то неизвестный нам естественный резонанс и как блокировку его следствия. Я отвечаю НЕТ !. За всю историю радиоэлектроники и изученных электрических схем было невозможно не обнаружить и не зафиксировать такой электромагнитный резонанс.4
С точки зрения местной среды — все осталось, как было раньше. Он, как и прежде, имеет электромагнитное сопротивление, всегда пытаясь уравновесить созданный локальный градиент от напряжения, «упругости». В развитии этой мысли возникает вопрос. Есть ли естественный резонанс окружающей среды. В смысле, могли бы мы случайно, используя нашу электрическую схему, найти какой-то неизвестный нам естественный резонанс и как блокировку его следствия. Я отвечаю НЕТ !. За всю историю радиоэлектроники и изученных электрических схем было невозможно не обнаружить и не зафиксировать такой электромагнитный резонанс.4
Но есть небольшая открытость. Это объем окружающей среды, ограниченный сторонами формы материального объекта, где электромагнитные волны могут отражаться обратно. В этом объеме мы можем наблюдать так называемый объемный резонанс, вызванный не только процессом индукции, но и потоком электромагнитного излучения. Но это вопрос, относящийся к фонду «WSG», и мы не будем вдаваться в него.
V.
ДВОЙНОЙ РЕЗОНАНС В СРЕДЕ
Что я буду делать дальше.Я удаляю емкость в схеме и на ее месте подключаюсь к генератору синусоидальной волны катушки, предпочтительно с небольшим коэффициентом нелинейного изменения, и исследую ту же локальную среду.
Местный — потому что он создается законом уменьшения мощности электромагнитной индукции в зависимости от расстояния от ее источника. И слава богу, иначе у нас был бы постоянный резонанс. Чтобы контролировать резонанс в окружающей среде, я помещаю небольшую индукционную катушку поверх первой и подключаю ее к осциллографу.
5
Что происходит? Мы можем видеть ту же самую резонансную среду, только намного лучше, потому что генератор, в отсутствие емкости, не только толкает окружающую среду в одном направлении, но и втягивает ее в другом. Я вижу оптимальную мощность колеблющегося экологического процесса. Я сразу думаю о добавлении небольшого потребителя, использующего эту энергию … Для этого я добавляю в катушку лампочку накаливания и вижу, что она слегка горит. Но в этот момент я вижу, что мой генератор начал извлекать из сети мощность, равную мощности лампы накаливания.В то же время резонанс в локальной среде начал затягиваться, и я понял, что в среде локального резонанса появился, по утверждению Н. Тесла, трение! Все мои добавления, созданные в схеме, создавали простой трансформатор, и моя свободная энергия была нарушена и исчезла. Со стыдом осознаю, что нарушено самое главное — резонанс местной среды !!! Свободная энергия может быть извлечена из резонансной среды только при условии, что она не нарушит резонанс !!! Теперь, как мы это делаем ?? !!
Но в этот момент я вижу, что мой генератор начал извлекать из сети мощность, равную мощности лампы накаливания.В то же время резонанс в локальной среде начал затягиваться, и я понял, что в среде локального резонанса появился, по утверждению Н. Тесла, трение! Все мои добавления, созданные в схеме, создавали простой трансформатор, и моя свободная энергия была нарушена и исчезла. Со стыдом осознаю, что нарушено самое главное — резонанс местной среды !!! Свободная энергия может быть извлечена из резонансной среды только при условии, что она не нарушит резонанс !!! Теперь, как мы это делаем ?? !!
VI.
ЭВОЛЮЦИЯ DIEA …
В этом случае нам на помощь приходит Ритмодинамика У. Иванова. Иванов описывает следующее развитие. Когда в единой энергосистеме Украины, унаследованной от бывшего СССР, частота 6
в Гц была уменьшена, произошел значительный дисбаланс, при котором электроэнергия передавалась из России Украине. Я назвал это Хосляцким резонансом. Именно это и важно для нас из всей досягаемости Rithmodinamic.
Интересно, знал ли об этом Капанадзе или случайно столкнулся с этим процессом.Кроме того, очень интересно, знал ли Альфред М. Хаббард (генератор Хаббарда) об этом свойстве потока энергии или также случайно столкнулся с ним. Здесь раскрывается ответ на наш вопрос: как извлечь свободную энергию из резонанса окружающей среды, окружающей среды !, а не из электрической цепи!
VII.
ВТОРОЙ СМЕШАННЫЙ РЕЗОНАНС ЛОКАЛЬНОЙ СРЕДЫ
Вот почему, используя аланлогию для локального резонанса смешанной частоты, извлеченного из общей более крупной, «правильной» резонансной энергетической системы, где существует скрытая свободная энергия, мы можем ограничиться поиском для параметров второй индукционной катушки и ее резонанса, созданного для извлечения энергии из уже созданного резонанса в локальной среде первой индукционной катушки.1. Объем энергии, извлеченный из второй индукции, должен быть величиной или 10-1 и даже больше, меньше, чем объемная энергия, созданная в первой индукции.
2. Как следствие этого, магнитное поле второй индукционной катушки должно быть намного меньше, чем магнитное поле первой индукционной катушки. 3. Из-за требований к магнитному полю физические параметры и индукционная емкость Генри второй индукционной катушки должны быть меньше, чем у первой. 4. Отвод мощности от второй индукционной катушки — это следствие разницы между резонансной частотой первой и второй индукционных катушек 7
!!!
Может ли случиться так, что частота второй индукционной катушки резко изменится таким образом, чтобы привлечь всю энергию из локального резонанса? Конечно может !!! В патенте Капанадзе об этом есть описание:
Первый регулятор частоты (7) стабилизирует созданную высокую частоту в соответствии с потребностью и наводит порядок в ней, не оказывая вредного воздействия на выходной контур.
По-видимому, упомянутое выше беспокоило его и, кажется, не только … Можно сказать, что извлечение энергии из резонанса окружающей среды также создает резонанс, но его частота находится в диапазоне Гц и десятых долях Гц, отклоняется от частоты резонанса, создаваемого первой катушкой. Какой должна быть конструкция электросхемы и какой должна быть разность частот для выполнения этих условий — определим позже.
Какой должна быть конструкция электросхемы и какой должна быть разность частот для выполнения этих условий — определим позже.
VIII.
ТРЕТИЙ РЕЗОНАНС — РЕЗОНАНС ЗАРЯДА
Завершив все вышеперечисленное и добавив лампочку накаливания ко второй катушке, все возвращается, как было раньше — мы получаем тот же базовый простой трансформатор, и с ним неприятное ощущение, что без записанная экспериментальная презентация Тариела Капанадзе, мы могли бы прекратить наши усилия в этом направлении.
8
Но мы продолжаем развивать нашу мысль. Кто нам сказал, что если мы добавим лампочку накаливания ко второй катушке, мы не изменим, не ухудшим условия резонанса в окружающей среде? Даже если вторая катушка имеет резонансную частоту, настроенную на соответствующую резонансную частоту первой индукционной катушки. Понятно, в чем проблема, она исчезнет, потому что второй резонанс из «плавно пульсирующего» переходит в фрикционный! Та же лампочка, которая использовалась раньше, когда мы подключались параллельно к оцицилоскопу. Тогда как мы можем это сделать? Как убрать это трение? Собственно, снимать его не нужно! Что такое электрический ток?
Тогда как мы можем это сделать? Как убрать это трение? Собственно, снимать его не нужно! Что такое электрический ток?
По данным института милиции: Главный проессор показывает движение тока по электрической схеме: от плюсовой розетки источника питания, от лампы накаливания или другой схемы, а затем на землю, тыква — источник энергии Итак, что такое электрический ток? Это только поток заряженных частиц, электронов через электрическую цепь? Лучшее определение, по-видимому, дано Горовицем и Хиллом, автором самой продаваемой книги «Искусство электроники».Электрический ток — это скорость движения электрического заряда в точке!
Итак, перед нами проблема: зажечь лампочку накаливания. Как мы можем это сделать?
1. Подайте разность потенциалов от источника постоянного тока, например аккумулятора. 2. Подайте переменное напряжение от электросети. 3. Но мы можем сделать это другим способом: поместите лампочку накаливания в катушку, подключенную к высокочастотной цепи, и лампочка загорится
без применения
любой разности потенциалов
! В последнем случае нет электрического тока, идущего к
, потенциал
!
наземный источник
В этом случае заряды, электроны, будут биться с высокой частотой и будут составлять 9
, нарушая кристаллическую сетку металла. Это хорошо. Если мы добавим в схему еще одну лампочку накаливания, взяв кабели от лампочки, которая находится в индукторе, и подключим их ко второй лампочке, расположенной за пределами индуктора. Вторая лампочка не загорится, а просто нагреется. Вторая лампочка не загорится, потому что мощности отскакивающих заряженных частиц недостаточно для создания отскока заряженных частиц в небольшом объеме гирлянды второй лампы накаливания. Мы снова видим точный процесс трения.Но если мы подключим лампочку
Это хорошо. Если мы добавим в схему еще одну лампочку накаливания, взяв кабели от лампочки, которая находится в индукторе, и подключим их ко второй лампочке, расположенной за пределами индуктора. Вторая лампочка не загорится, а просто нагреется. Вторая лампочка не загорится, потому что мощности отскакивающих заряженных частиц недостаточно для создания отскока заряженных частиц в небольшом объеме гирлянды второй лампы накаливания. Мы снова видим точный процесс трения.Но если мы подключим лампочку
, одну из клемм ко второй лампе накаливания
,, к любому большому железному объекту, а еще лучше, к земле, как это делает г-н Капанадзе в фильме
, то картина сразу же изменится. У нас есть большой свободный источник заряженных частиц, это наша Земля. Нам не нужно вдавливать или выталкивать это ограниченное количество заряженных частиц, находящихся в маленькой вольфрамовой катушке лампочки. Мы создаем на свободном конце противоположный потенциал, достигающий сотен вольт, точно так же, как он создается в небольшой вольфрамовой катушке, расположенной в высокочастотной катушке индуктивности.Самое главное, что мы искали, это создать — когда у нас есть неограниченный объем заряженных частиц, исходящих от большого объекта —
резонанса заряженных частиц !!!
,
в массе толстой медной проволоки, «созданной» посредством первой и второй индукций. Нет электрического тока в обычном понимании этого слова! Есть только «подпрыгивание» заряженных частиц в массе толстой медной проволоки! И с учетом этих выводов нагрузка прилагается к зеленому ящику.Вот почему «подпрыгивание» электронов в массе толстого провода, второй конец которого абсолютно необходимо соединить с заземлением, может возникнуть, когда к нему через нагрузку присоединен только один конец второй индукционной катушки! !!
10
Самое главное, чтобы не было препятствий при трении, для возникновения третьего резонанса — резонанса заряженных частиц. С вашего позволения gentelemnt, еще раз повторяю: электрического тока нет, как это принято в обиходе, только трение.Вот почему розетка, обозначенная зеленым прямоугольником, основана на высоких частотах! Самая оптимальная нагрузка в этом случае — это активная нагрузка, ведь именно такова нагрузка лампы накаливания. Этот факт сам по себе создает принципиально новый способ работы с активной нагрузкой в электродвигателях и т.п. Но лучшее из реактивных нагрузок в том, что в этом типе схемы мы можем создать дополнительный резонанс, резонанс, специфичный для извлеченной нагрузки! Мы задаем себе следующий актуальный вопрос: как подключить к такой электрической цепи широко распространенный электродвигатель переменного тока, работающий на частоте 50 Гц? Для этого Mr.Капанадзе использует обычный амплитудный модулятор на 50 Гц, обозначенный на схеме патента номером 11.
IX.
ТАК, ЧТО У НАС?
Мы рассмотрели состояние деревьев, резонансные системы деревьев, которые должны быть на месте, чтобы получать бесплатную энергию.
11
1. Высоковольтная высокочастотная резонансная среда, созданная в первой индукционной катушке. 2. Высокочастотный резонанс во второй индукционной катушке, но этот резонанс должен быть низким напряжением и иметь другую частоту и номинальную мощность от резонанса в первой индукционной катушке.3. Гальванический резонанс заряженных частиц в корпусе толстой медной проволоки, заземленной на одном конце для приема неограниченного количества заряженных частиц, чтобы противодействовать процессу трения в гальваническом резонансе. Потому что в разработанном устройстве все влияет друг на друга, и процесс трения гальванического резонанса влияет на резонанс в первой индукционной катушке. 4. Особый, отличный от традиционного, способ подключения рабочей нагрузки. Очень интересно настроить соотношение мощности резонанса в первой и второй катушках.Если учесть, что искровой промежуток в зеленом квадрате составляет около 1–1,5 мм, напряжение должно быть в пределах 2 × 103 вольт. Частота, приложенная к искровому промежутку, должна быть около — 150 × 103 — 200 × 103 Гц. Получается расчетный ток:
5000Вт 220В
= 23А, именно это амперметр
показывает в фильме. Получается, что мощность в первой цепи в 10 раз больше и составляет 50 × 103 Вт.
Это правда? Здесь мы имеем дело с резонансом.Сделаем расчеты другим способом. В первой катушке 2000 × 2000 = 4 × 106 Во второй катушке 220 × 220 = 48,4 × 103 Соотношение
4 × 106 48,4 × 103
≈ 82 или руй
100 раз
.
Речь идет о 2-х величинах. Так что это что-то. Итак, в условиях локального резонанса в первой индукционной катушке присутствует пульсирующая мощность 500 × 103 Вт, а мы извлекаем довольно небольшую — 5 × 103 Вт. На принципиальном уровне это может быть шум, так что «военное формирование (100 солдат) не почувствует пропавшего 1 человека 5 × 103 Вт.Теперь посмотрим на частоту. Если в «резонансе Хохлатского» декремент частоты в энергосистеме составлял 0,5 Гц, это 1% от 50 Гц. 1% от 200 × 103 Гц = 2 × 103 Гц
Получается, что частота второй катушки в 100 раз меньше, то есть в области 2 × 103 Гц. Кажется, что существует денитная зависимость между мощностью и частотой.
Но это для будущего расследования. Похоже, что это намного лучше, чем пытаться получить забавный резонанс во второй индукционной катушке в Гц или даже в десятках Гц, как я делал это раньше.
X.
ЗАКЛЮЧЕНИЕ
Разработаны основные принципы решения данной проблемы. Есть некоторые проблемы с теорией контуров, но я объяснил их ранее, чтобы это не опровергало мою точку зрения. В заключение я показываю схему, подтверждающую объяснение Капанадзе, что «конденсатор (2) используется для извлечения …», и здесь наглядно показано его использование в качестве переключателя.
13
Итак, теперь, пожалуйста, выскажите свое мнение, Gentelmen! P.S В принципе, господа, здесь как-то странно взаимосвязано измерение времени и передача энергии времени, и это причина, которая и спровоцировала мое исследование вопроса.Но это тема для более серьезного разговора.
14
недавних сильных вспышек рентгеновского излучения 1ES 1959 + 650 с, возможно, менее эффективным стохастическим ускорением | Ежемесячные уведомления Королевского астрономического общества: письма
Аннотация
Мы представляем вспышку в рентгеновских лучах 1ES 1959 + 650 в августе – январе 2016 г., которая была самой мощной и продолжительной за 10,75 лет с начала ее мониторинга с помощью рентгеновского телескопа на борту Swift .Новая максимальная историческая скорость счета 0,3–10 кэВ была зарегистрирована три раза, что делает этот объект третьим источником BL Lacertae, превышающим уровень в 20 отсчетов s −1 . Эта эпоха наряду с общей переменностью в 5.7 раза характеризовалась быстрыми рентгеновскими вспышками в 2.0–3.1 раза, сопровождавшимися экстремальной спектральной изменчивостью. Источник также показывает одновременную вспышечную активность в оптическом — УФ диапазонах и 0,3–100 ГэВ, хотя также обнаруживается быстрая вспышка в γ-квантах без значительных оптических и рентгеновских аналогов.В отличие от рентгеновских вспышек в предыдущие годы, стохастическое ускорение кажется менее важным для электронов, ответственных за производство рентгеновского излучения во время этой вспышки, что ставит под сомнение ранее высказанное предположение, что электроны в струях обнаруженного ТэВ BL Lacertae объекты должны подвергаться эффективному стохастическому ускорению, что приводит к меньшей кривизне рентгеновского спектра.
1 ВВЕДЕНИЕ
Объекты BL Lacertae (BLLs) широко считаются активными галактическими ядрами (AGN) с релятивистской струей, тесно выровненной с лучом зрения, что успешно объясняет их характерные особенности, такие как нетепловое излучение континуума по всему спектру и сильное переменность потока во всех спектральных диапазонах (Massaro, Paggi & Cavaliere 2011a).Спектральные распределения энергии (SED) BLLs показывают присутствие двух широких компонентов. Принято считать, что более низкочастотное излучение связано с синхротронным излучением, испускаемым релятивистскими электронами в струе, в то время как обратное комптоновское (IC) рассеяние синхротронных фотонов на той же электронной населенности должно вызывать высокочастотный удар (так называемый синхротронный самотек). -Механизм Комптона, SSC), хотя также предлагаются альтернативные сценарии (см. Krawczynski et al. 2004). Мы можем сделать выводы о применимости этих моделей, изучая характер переменности потока в различных спектральных диапазонах и их взаимные корреляции, которые, в свою очередь, дают очень важные подсказки о физике, структуре и динамике излучения BLL. зона.
Здесь мы сообщаем о сильной вспышке рентгеновского излучения обнаруженного ТэВ BLL 1ES 1959 + 650 ( z = 0,047), наблюдаемого с помощью рентгеновского телескопа (XRT; Берроуз и др., 2005) на борту Swift. (Герельс и др., 2004) в течение августа 2015 г. — января 2016 г., что было значительно сильнее и продолжительнее по сравнению с сообщениями (Капанадзе и др., 2016, далее K16), Алиу и др. (2013), Tagliaferri et al. (2008) и др. По рентгеновским наблюдениям 2000–14 гг.
2 НАБЛЮДЕНИЯ И ОБРАБОТКА ДАННЫХ
Данные XRT обрабатывались с помощью пакета xrtdas, разработанного в Центре научных данных ASI (ASDC) и распространяемого HEASARC в пакете heasoft (v. 6.16). Они были уменьшены, откалиброваны и очищены с помощью скрипта xrtpipeline с использованием стандартных критериев фильтрации и файлов калибровки Swift caldb (20150721). Извлечение кривых блеска и спектров 0,3–10 кэВ из наблюдений Windowed Timing (WT) и Photon Count (PC), а также их корректировка по различным эффектам выполнялись таким же образом, как подробно описано в K16.Фотометрия изображений с коррекцией неба, полученных во всех диапазонах UVOT, была выполнена с помощью программы heasoft task uvotsource и файлов калибровки, включенных в caldb. Измерения проводились с использованием апертуры источника радиуса 5 угловых секунд для диапазонов V, B, U и радиуса 10 угловых секунд — для диапазонов UVW1 , UVM2 , UVW2 , чтобы должным образом учесть более широкие PSF. Затем величины были скорректированы с учетом галактического поглощения и преобразованы в линейные потоки (в мЯн) в соответствии с рецептом, приведенным в K16.
Мы извлекли потоки фотонов 300 МэВ – 100 ГэВ из наблюдений Fermi -LAT. События диффузного класса использовались из интересующей области (ROI) с радиусом 10 градусов с центром в координатах 1ES 1959 + 650 и обрабатывались с помощью пакета Fermi Science Tools (версия v10r0p5) с функцией отклика прибора P8R2_V6. Сделаны разрезы по зенитному углу (> 100 градусов) и углу качания КА (> 52 градуса). Фоновая модель, включающая все источники гамма-излучения из 4-летнего каталога точечных источников Fermi –LAT (3FGL, Acero et al.2015) в пределах 20 градусов от 1ЭС 1959 + 650. Спектральные параметры источников в пределах области интереса остаются свободными во время процесса минимизации, в то время как параметры вне этого диапазона удерживаются фиксированными на значениях каталога. Галактическое и внегалактическое диффузное гамма-излучение, а также остаточный инструментальный фон включены с использованием рекомендованных файлов моделей.
В период со 2 августа по 10 декабря 2015 г. источник наблюдался при энергиях ТэВ в течение 151,8 ч (распределен в течение 96 ночей) на Первом черенковском телескопе G-APD (FACT; Anderhub et al.2013), расположенный в Observatorio del Roque de los Muchachos (Ла-Пальма, Испания). Сотрудничество FACT предоставляет результаты предварительного быстрого анализа на http://www.fact-project.org/monitoring. Эти кривые блеска за вычетом фона не корректируются с учетом эффекта изменения энергетического порога с изменением зенитного расстояния и окружающего освещения, а также не производится выбор качества данных (Дорнер и др., 2015). Во время фазы повышенного потока была обнаружена 1ES 1959 + 650 с более чем 3σ в три ночи 18/19 октября 2015 г. (MJD 57313.8), 15/16 ноября (57341,8 MJD) и 20/21 ноября (57346,8 MJD).
3 РЕЗУЛЬТАТЫ, ОБСУЖДЕНИЕ И ВЫВОДЫ
3.1 Сроки
Источник наблюдался 52 раза с помощью XRT в период с 1 августа 2015 года по 19 января 2016 года, в основном в рамках инициированных нами наблюдений за Целевыми возможностями (ToO). Информация о каждом наведении и результаты измерений представлены в Таблице 1. 1 Результаты наблюдений UVOT представлены в Таблице 2, где уменьшенные величины и соответствующие потоки представлены для каждого диапазона.
Таблица 1.Выдержка из сводки наблюдений Swift –XRT. Столбцы (3) и (4) показывают время экспозиции (в секундах) и среднюю скорость счета 0,3–10 кэВ с ошибкой в скобках для данного ObsID (в отсчетах с –1 ) соответственно; Столбец (5) — значение χ 2 с соответствующей d.o.f для конкретного наблюдения.
| ObsID . | Обс. Начало — Конец (UTC) . | Exp. . | Скорость счета . | χ 2 / д.о.ф. . |
|---|---|---|---|---|
| (1) . | (2) . | (3) . | (4) . | (5) . |
| 00035025149 | 2015-08-01 22:20:58 08-01 23:22:22 | 550 | 10,08 (0,26) | 9.9/8 |
| 00035025151 | 2015-08-15 11:49:58 08-15 14:10:17 | 959 | 7,50 (0,10) | 33,6 / 14 |
| ObsID . | Обс. Начало — Конец (UTC) . | Exp. . | Скорость счета . | χ 2 / д.о.ф. . |
|---|---|---|---|---|
| (1) . | (2) . | (3) . | (4) . | (5) . |
| 00035025149 | 2015-08-01 22:20:58 08-01 23:22:22 | 550 | 10,08 (0,26) | 9,9 / 8 |
| 00035025151 | 2015 08-15 11:49:58 08-15 14:10:17 | 959 | 7,50 (0,10) | 33,6 / 14 |
Выдержка из резюме наблюдений Swift –XRT .Столбцы (3) и (4) показывают время экспозиции (в секундах) и среднюю скорость счета 0,3–10 кэВ с ошибкой в скобках для данного ObsID (в отсчетах с –1 ) соответственно; Столбец (5) — значение χ 2 с соответствующей d.o.f для конкретного наблюдения.
| ObsID . | Обс. Начало — Конец (UTC) . | Exp. . | Скорость счета . | χ 2 / д.из. . |
|---|---|---|---|---|
| (1) . | (2) . | (3) . | (4) . | (5) . |
| 00035025149 | 2015-08-01 22:20:58 08-01 23:22:22 | 550 | 10,08 (0,26) | 9,9 / 8 |
| 00035025151 | 2015 08-15 11:49:58 08-15 14:10:17 | 959 | 7.50 (0,10) | 33,6 / 14 |
| ObsID . | Обс. Начало — Конец (UTC) . | Exp. . | Скорость счета . | χ 2 / д.о.ф. . |
|---|---|---|---|---|
| (1) . | (2) . | (3) . | (4) . | (5) . |
| 00035025149 | 2015-08-01 22:20:58 08-01 23:22:22 | 550 | 10,08 (0,26) | 9,9 / 8 |
| 00035025151 | 2015 08-15 11:49:58 08-15 14:10:17 | 959 | 7,50 (0,10) | 33,6 / 14 |
Выдержка из результатов наблюдений Swift –UVOT. Значения потока в каждой полосе даны в мЯн.
| . | В . | B . | U . | UVW1 . | УВМ2 . | UVW2 . | ||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| ObsId | Mag. | Флюс | Маг. | Флюс | Маг. | Флюс | Маг. | Флюс | Маг. | Флюс | Маг. | Флюс |
| 35025149 | 14,54 (0,06) | 5,60 (0,30) | 14,94 (0,04) | 4,29 (0,17) | 14,06 (0,04) | 1439 (0,139) | 2,21 (0,12) | 13,77 (0,06) | 2,38 (0,18) | 13,88 (0,05) | 2,07 (0,10) | |
| 35025151 | 14.42 (0,06) | 6,25 (0,36) | 14,74 (0,04) | 5,15 (0,21) | 13,94 (0,05) | 3,84 (0,18) | 13,79 (0,06) | 2,73 (0,15) 13,6397 | 0,06)2,61 (0,14) | 13,57 (0,05) | 2,75 (0,13) | |
| . | В . | B . | U . | UVW1 . | УВМ2 . | UVW2 . | ||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| ObsId | Mag. | Флюс | Маг. | Флюс | Маг. | Флюс | Маг. | Флюс | Маг. | Флюс | Маг. | Флюс |
| 35025149 | 14,54 (0,06) | 5.60 (0,30) | 14,94 (0,04) | 4,29 (0,17) | 14,06 (0,04) | 3,44 (0,16) | 14,02 (0,05) | 2,21 (0,12) | 13,77 (0,06) | 0,18)13,88 (0,05) | 2,07 (0,10) | |
| 35025151 | 14,42 (0,06) | 6,25 (0,36) | 14,74 (0,04) | 5,15 (0,05) | 3,84 (0,18) | 13,79 (0,06) | 2,73 (0.15) | 13,67 (0,06) | 2,61 (0,14) | 13,57 (0,05) | 2,75 (0,13) | |
Выдержка из результатов наблюдений Swift –UVOT. Значения потока в каждой полосе даны в мЯн.
| . | В . | B . | U . | UVW1 . | УВМ2 . | UVW2 . | ||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| ObsId | Mag. | Флюс | Маг. | Флюс | Маг. | Флюс | Маг. | Флюс | Маг. | Флюс | Маг. | Flux |
| 35025149 | 14,54 (0,06) | 5,60 (0,30) | 14,94 (0,04) | 4.29 (0,17) | 14,06 (0,04) | 3,44 (0,16) | 14,02 (0,05) | 2,21 (0,12) | 13,77 (0,06) | 2,38 (0,18) | 13,88 (0,05) | 0,10)|
| 35025151 | 14,42 (0,06) | 6,25 (0,36) | 14,74 (0,04) | 5,15 (0,21) | 13,94 (0,05) | 3,84 (0,18) | 3,84 (0,18) | 2,73 (0,15) | 13,67 (0,06) | 2,61 (0.14) | 13,57 (0,05) | 2,75 (0,13) |
| . | В . | B . | U . | UVW1 . | УВМ2 . | UVW2 . | ||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| ObsId | Mag. | Флюс | Маг. | Флюс | Маг. | Флюс | Маг. | Флюс | Маг. | Флюс | Маг. | Флюс |
| 35025149 | 14,54 (0,06) | 5,60 (0,30) | 14,94 (0,04) | 4,29 (0,17) | 14,06 (0,04) | 1439 (0,139) | 2,21 (0,12) | 13,77 (0,06) | 2,38 (0,18) | 13,88 (0,05) | 2.07 (0,10) | |
| 35025151 | 14,42 (0,06) | 6,25 (0,36) | 14,74 (0,04) | 5,15 (0,21) | 13,94 (0,05) | 3,84 (0,14) | 3,84 (0,18) ) | 2,73 (0,15) | 13,67 (0,06) | 2,61 (0,14) | 13,57 (0,05) | 2,75 (0,13) |
Корреляции между различными спектральными параметрами и многополосными потоками.
| Кол-во . | r . | п. . |
|---|---|---|
| F 0,3−2 кэВ и F 2-10 кэВ | 0,74 (0,05) | 7,14 × 10 −13 | −0,40 (0,11) | 1,35 × 10 −4 |
| HR и F 0.3-10 кэВ | 0,55 (0,08) | 2,60 × 10 −6 |
| E p и F 0,3-10 кэВ | 0,45 90 7,5 (0,12) 10 −5 |
| Количество . | r . | п. . | |||
|---|---|---|---|---|---|
| F 0,3−2 кэВ и F 2-10 кэВ | 0.74 (0,05) | 7,14 × 10 −13 | |||
| a и F 0,3−10 кэВ | −0,40 (0,11) | 1,35 × 10 365 −4 | HR и F 0,3-10 кэВ | 0,55 (0,08) | 2,60 × 10 −6 |
| E p и F 4 0,3-1039 кэВ 0,45 (0,12) | 7.51 × 10 −5 |
Корреляции между различными спектральными параметрами и многополосными потоками.
| Кол-во . | r . | п. . |
|---|---|---|
| F 0,3−2 кэВ и F 2-10 кэВ | 0,74 (0,05) | 7,14 × 10 −13 | -0,40 (0,11) | 1,35 × 10 -4 |
| HR и F 0,3-10 кэВ | 0,55 (0,08) | 2,94 −6 |
| E p и F 0,3−10 кэВ | 0,45 (0,12) | 7,51 × 10 −5 |
| ObsId . | a . | b . | E p . | К . | χ 2 / д.о.ф. . | log F 0,3−2 кэВ . | log F 2-10 кэВ . | log F 0,3-10 кэВ . | HR . |
|---|---|---|---|---|---|---|---|---|---|
| (1) . | (2) . | (3) . | (4) . | (5) . | (6) . | (7) . | (8) . | (9) . | (10) . |
| 35025149 | 1,95 (0,09) | 0,77 (0,19) | 1.08 (0,10) | 6,42 (0,14) | 48,2 / 54 | −9,986 (0,021) | −10,288 (0,039) | −9.809 (0,021) | 0,499 (0,051) |
| 2,09 (0,06) | 0,51 (0,13) | 0,82 (0,10) | 6,51 (0,14) | 92,5 / 86 | -9,832 (0,016) | -10,15 (0,029) | -9,662 | 0,481 (0,037)
| ObsId . | a . | b . | E p . | К . | χ 2 / д.о.ф. . | log F 0,3−2 кэВ . | log F 2-10 кэВ . | log F 0,3-10 кэВ . | HR . |
|---|---|---|---|---|---|---|---|---|---|
| (1) . | (2) . | (3) . | (4) . | (5) . | (6) . | (7) . | (8) . | (9) . | (10) . |
| 35025149 | 1,95 (0,09) | 0,77 (0,19) | 1.08 (0,10) | 6,42 (0,14) | 48,2 / 54 | −9,986 (0,021) | −10,288 (0,039) | −9.809 (0,021) | 0,499 (0,051) |
| 35025151 | 2,09 (0,06) | 0,51 (0,13) | 0,82 (0,10) | 6,51 (0,14) | 92,5 / 86 | -9,832 (0,016) | -10,15 (0,029) | -9,662 | 0,481 (0,037)
Выдержка из краткого обзора спектрального анализа XRT с моделью LP.Значения E p (столбец 4) даны в кэВ; параметр K (столбец 5) задается в единицах 10 −2 ; непоглощенные потоки 0,3–2, 2–10 и 0,3–10 кэВ (столбцы 7–9) — в эрг см –2 с –1 .
| ObsId . | a . | b . | E p . | К . | χ 2 / д.о.ф. . | log F 0,3−2 кэВ . | log F 2-10 кэВ . | log F 0,3-10 кэВ . | HR . |
|---|---|---|---|---|---|---|---|---|---|
| (1) . | (2) . | (3) . | (4) . | (5) . | (6) . | (7) . | (8) . | (9) . | (10) . |
| 35025149 | 1,95 (0,09) | 0,77 (0,19) | 1,08 (0,10) | 6,42 (0,14) | 48,2 / 54 | -9,988 (0,021) 9,2 | −9.809 (0,021) | 0,499 (0,051) | |
| 35025151 Орбита1 | 2.09 (0,06) | 0,51 (0,13) | 0,82 (0,10) | 6,51 (0,14) | 92,5 / 86 | -9,832 (0,016) | -10,15 (0,029) | -9,662 (0,0154) | 0,481 (0,037)
| ObsId . | a . | b . | E p . | К . | χ 2 / д.о.ф. . | log F 0,3−2 кэВ . | log F 2-10 кэВ . | log F 0,3-10 кэВ . | HR . |
|---|---|---|---|---|---|---|---|---|---|
| (1) . | (2) . | (3) . | (4) . | (5) . | (6) . | (7) . | (8) . | (9) . | (10) . |
| 35025149 | 1,95 (0,09) | 0,77 (0,19) | 1,08 (0,10) | 6,42 (0,14) | 48,2 / 54 | -9,988 (0,021) 9,2 | −9.809 (0,021) | 0,499 (0,051) | |
| 35025151 Орбита1 | 2.09 (0,06) | 0,51 (0,13) | 0,82 (0,10) | 6,51 (0,14) | 92,5 / 86 | -9,832 (0,016) | -10,15 (0,029) | -9,662 (0,0154) | 0,481 (0,037)
Распределение различных спектральных параметров.
| . | 2015 август – 2016 январь . | 2005–14 . | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Пар. | Мин. | Макс. | Пик | σ 2 | Мин. | Макс. | Пик | σ 2 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| b | 0,28 | 0,98 | 0,49 | 0,021 | 0,18 | 0,67 | 9039 | 2,09 | 1,74 | 0,028 | 1.76 | 2,37 | 2,02 | 0,016 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| HR | 0,481 | 1,496 | 0,845 | 0,043 | 0,317 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 0,317 | 0,94 184 | 0,317 | 0,94 . | 2015 август – 2016 январь . | 2005–14 . | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Пар. | Мин. | Макс. | Пик | σ 2 | Мин. | Макс. | Пик | σ 2 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| b | 0,28 | 0,98 | 0,49 | 0,021 | 0,18 | 0,67 | 9039 | 2,09 | 1,74 | 0,028 | 1,76 | 2.37 | 2,02 | 0,016 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| HR | 0,481 | 1,496 | 0,845 | 0,043 | 0,317 | 0,043 | 0,317 | 0,043 | 0,317 | 0,918 | 0,550 9017 | 0,550 9039 параметры.
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||